Перепончатокрылые. Перепончатокрылые насекомые фото описание
Перепончатокрылые – многочисленный и наиболее развитый отряд насекомых.
Отличительным признаком этого семейства является то, что при наличии двух пар перепончатых крыльев, задние не функционируют во время полета.
Существуют виды бескрылых перепончатых насекомых. Осы, муравьи, пчелы, шмели – самые известные представители семейства перепончатокрылых насекомых.
Их значение в экосистеме нашей планеты огромно и является своего рода лакмусовой бумажкой определяющей ее экологическое благополучие.
К многочисленному отряду перепончатокрылых относятся как самые маленькие насекомые, так и самые гигантские. Длина тела самцов паразитических наездников около 0,139 мм. Наиболее крупные перепончатокрылые – тропические и дорожные осы, сколии и некоторые виды наездников имеют огромные размеры от 4 до 6 см.
перепончатокрылые.
Иногда длина яйцеклада значительно увеличивает размеры некоторых насекомых. Все части тела четко разграничены. Голова имеет сложные и простые глаза. Преобладают ротовые органы грызущего типа. Используется не только для приема пищи, но и при строительстве жилья, пережевывания пищи для личинок, как боевой арсенал в битвах.
Известны случаи прогрызания свинцовых листов отдельными видами перепончатокрылых. Членистые сегменты груди составляют единое целое. Разные виды имеют членистое брюшко разнообразных форм. Большинство взрослых особей самок имеют в задней части брюшка жало или сверло, у самцов там находятся органы совокупления.
перепончатокрылые. шмель фото
Ногам свойственные разнообразные функции – ходильные, хватательные, копательные. Две пары крыльев прозрачные или слабо окрашенные. Некоторые виды бескрылые, например рабочие муравьи.
Для экологического равновесия важны самые разные представители семейства перепончатокрылых. Пчелы, шмели, муравьи и некоторые другие виды выполняют важнейшую функцию для сохранения флоры – являются опылителями.
Пчелы, шмели, муравьи и некоторые другие виды выполняют важнейшую функцию для сохранения флоры – являются опылителями.
Хищники, например осы или муравьи и насекомые-паразиты, например наездники, играют роль естественных регуляторов популяций, являющихся для них пищей. Перепончатокрылые, например пилильщики, питающиеся растительной пищей, съедают существенную часть ежегодного прироста.
перепончатокрылые. оса фото
Способность отдельных видов муравьев переносить семена некоторых растений получила специальный термин мирмекохория. Они «рассаживают” около 1% от всех растений нашей планеты. Благодаря этому в лесах и на лугах сохраняются многие редкие виды растений. Среди перепончатокрылых каннибализм обычное и широко распространенное явление.
Размножаются откладыванием яиц, используя яйцеклад или свёрла. Яйца откладывают в специально приготовленные жилища в дереве, земле – пчелы, осы, муравьи.
Иногда яйца откладывают в сочные части растения, в гнезда других насекомых или прямо на животных.
Так размножаются некоторые наездники, пчёлы-кукушки, блестянки, орехотворки. Перепончатокрылые проходят полную стадию развития – яйцо, личинка, куколка, взрослая особь. Среда обитания Перепончатокрылые существуют повсюду – на земле и в почве, в воде, в воздухе и в других живых организмах.
перепончатокрылые. муравей фото
Некоторые виды строят специальные жилища, другие предпочитают вести паразитический образ. Пчелы и осы строят специальные гнезда и заботятся о потомстве, наполняя гнездо медом и цветочной пыльцой.
- Муравьи могут использовать тлю как «домашний скот”. Они ее выхаживают, пасут, охраняют и даже доят, получая для питания специфическую жидкость.
- У муравьев существует четкая иерархия и разделение обязанностей, за неисполнение которых муравья могут казнить и съесть.
- Некоторые виды шмелей специально разводят для использования на дачных участках в качестве опылителей растений.

35. Особенности строения, образа жизни, питание, многообразие отряда перепончатокрылых.
Тип: Членистоногие
Класс: Насекомые
Отряд: Перепончатокрылые
Перепончатокры́лые (Hymenoptera) – имеет две пары крыльев задние меньше передних, крылья с редкой сетью жилок, редко без жилок. на переднем крае заднего крыла расположен ряд крючковидных зацепок, входящих в соответствующую им складку на заднем крае переднего крыла, грызущие и лижущие или только грызущие ротовые органы и полное превращение. Размер варьирует от 0,2 мм до 135 мм.
Голова,
грудь и брюшко резко обособлены. Сложные
глаза почти всегда хорошо развиты,
между ними обыкновенно три расположенных
в виде треугольника простых глазка;
встречаются также формы, лишённые
глазков или совершенно слепые (например,
рабочие муравьи.
Усики
имеют от 3 до 60 члеников; они бывают
прямые или коленчатые. Ротовые органы
или чисто грызущего типа, или же нижняя
губа и нижние челюсти изменены в органы
принятия жидкой пищи (органы лизания).
Крылья всегда перепончатые, прозрачные
или окрашенные, с редкой сетью жилок,
иногда вовсе без них; первая пара крыльев
всегда длиннее задней. Ноги перепончатокрылых
ходильные. Брюшко состоит из 6—8 члеников,
с члеником, плотно соединённого с
заднегрудью, и видоизмененных члеников,
втянутых в задний конец брюшка и несущих
у самок жало или сверло, у самцов —
органы совокупления. Придатки заднего
конца брюшка (жала и сверла или яйцеклады)
состоят из пары щетинок, или стилетов,
и желобковатой части, состоящей из двух
отдельных или более или менее слитных
пластинок. При основании жала, которое
в спокойном состоянии всегда втянуто
внутрь брюшка, находится парная ядовитая
железа с резервуаром, в котором
скапливается яд. Пищеварительный канал
сильно развит.
Превращение перепончатокрылых полное. Личинки представляют весьма различное строение. У одних (пилильщиков) личинки живут свободно на листьях, по внешнему виду похожи на личинок (гусениц) бабочек (отсюда название — ложные гусеницы) и имеют 3 пары грудных и 6—8 пар брюшных ног. У рогохвостов (Siricidae) личинки живут в дереве или (Cephidae) в стеблях и ветвях растений и имеют 3 пары недоразвитых грудных ног. У остальных перепончатокрылых личинки живут в гнездах или внутри питательных веществ и бывают лишены ног.
Куколки перепончатокрылых всегда относятся к типу свободных куколок . яйца просто откладываются в питательные растительные вещества, например листья или древесину.
Источником
питания служат нектар и пыльца цветков,
сок растений. Для многих видов
перепончатокрылых паразитов характерно
питание гемолимфой хозяина. Самки
внутренних паразитов после откладки
яйца слизывают выступающую на поверхность
тела капельку гемолимфы.
К отряду перепончатокрылых принадлежит значительное число видов (различные авторы определяют примерную цифру их в 15—25 тысяч
Большой вред культурным зерновым злакам (ржи, пшенице, ячменю, овсу) причиняют стеблевые пилильщики (Cephidae), особенно хлебный (Cephus pygmaeus) и черный (Trachelus tabidus)
По отношению к человеку некоторые перепончатокрылые полезны прямо, доставляя ему полезные вещества (мёд, воск), другие — косвенно, истребляя вредных насекомых (наездники). Важное значение имеют многие из перепончатокрылых также в процессе опыления растений. Вредят перепончатокрылые ужалениями, поедая различные запасы и портя здания (муравьи), но главным образом вредны многие из них в сельскохозяйственном отношении, причиняя более или менее значительные опустошения среди культурных и лесных растений.
Наибольшее
значение перепончатокрылые имеют как
объекты, применяющиеся в биологической
борьбе с насекомыми-вредителями.
ПЕРЕПОНЧАТОКРЫЛЫЕПЕРЕПÓНЧАТОКРЫ́ЛЫЕ( Hymenoptera) ,один из крупнейших отрядов насекомых, включающий более 300 тыс. или муравьёв). Для П. характерно наличие (у большинства видов) двух пар крыльев с небогатым жилкованием. Задние крылья значительно меньше передних и в полете сцепляются с ними (чаще с помощью специальных крючочков) в функционально единые крыловые поверхности. У мелких форм число жилок на крыльях уменьшено, вплоть до их полного отсутствия. Ротовой аппарат большинства П. характеризуется наличием более или менее вытянутого хоботка (из слившихся нижней губы и нижних челюстей), а также грызущих верхних челюстей. Это позволяет насекомым потреблять и жидкую и твердую пищу, а также осуществлять сложную строительную деятельность. В грудном отделе тела нижняя часть переднегруди с причленяющимися к ней ногами и головой образует функционально единый аппарат, также способствующий развитию сложных форм строительного поведения. По строению брюшка П. делят на 2 подотряда – сидячебрюхих (Symphyta) с широким основанием брюшка, целиком сливающимся с заднегрудью, и стебельчатобрюхих (Apocrita), имеющих сильно суженное к основанию брюшко («висячее») или брюшко, вытянутое в основании в длинный и тонкий стебелёк («стебельчатое»). Такое строение брюшка обеспечивает его высокую подвижность, необходимую при оборонительном ужаливании, парализации добычи или откладке яиц. При этом у стебельчатобрюхих морфологически первый сегмент брюшка вошёл в состав груди, полностью с нею слившись. Конец брюшка у самок несёт яйцеклад или жало. П. – насекомые с полным превращением. Личинки почти всех видов имеют прядильные желёзы, выделяющие при окукливании паутину для кокона. Для представителей отряда характерно развитие самок из оплодотворённых диплоидных яиц, а самцов – из неоплодотворённых гаплоидных. В некоторых группах (например, пилильщиков, муравьёв) из неоплодотворённых яиц могут развиваться и самки. По строению брюшка П. делят на 2 подотряда – сидячебрюхих (Symphyta) с широким основанием брюшка, целиком сливающимся с заднегрудью, и стебельчатобрюхих (Apocrita), имеющих сильно суженное к основанию брюшко («висячее») или брюшко, вытянутое в основании в длинный и тонкий стебелёк («стебельчатое»). Такое строение брюшка обеспечивает его высокую подвижность, необходимую при оборонительном ужаливании, парализации добычи или откладке яиц. При этом у стебельчатобрюхих морфологически первый сегмент брюшка вошёл в состав груди, полностью с нею слившись. Конец брюшка у самок несёт яйцеклад или жало. П. – насекомые с полным превращением. Личинки почти всех видов имеют прядильные желёзы, выделяющие при окукливании паутину для кокона. Для представителей отряда характерно развитие самок из оплодотворённых диплоидных яиц, а самцов – из неоплодотворённых гаплоидных. В некоторых группах (например, пилильщиков, муравьёв) из неоплодотворённых яиц могут развиваться и самки.
|
 описанных видов. Будучи достаточно сходными по морфологии, они обнаруживают широчайшее разнообразие типов экологических приспособлений и поведенческих реакций, особенно в формах так называемого социального, или общественного поведения (например, у
описанных видов. Будучи достаточно сходными по морфологии, они обнаруживают широчайшее разнообразие типов экологических приспособлений и поведенческих реакций, особенно в формах так называемого социального, или общественного поведения (например, у
Перепончатокрылые (Hymenoptera) / Курс / Энтомология
Перепончатокрылые не имеют родственных связей с каким-либо другим отрядом крылатых насекомых, характеризуются грызущим или грызуще-лижущим ротовым аппаратом, двумя парами перепончатых крыльев и развитым яйцекладом (рис. 220, Б). По числу видов (90 000) они уступают лишь жукам и бабочкам, намного превосходят их по уровню организации и формам поведения. Наряду с двукрылыми они могут считаться самыми совершенными насекомыми и, видимо, отделились как самостоятельная группа на самых ранних этапах эволюции.
Рисунок 220. Бабочка (А) и перепончатокрылое (Б)
В пределах отряда обычно выделяют относительно примитивных сидячебрюхих (Symphyta) и более развитых стебельчатобрюхих (Apocrita), отличающихся наличием стебелька между грудным отделом и брюшком. Принимаемое ранее разделение стебельчатобрюхих на паразитических (Parasitica) и жалящих (Aculeata) сейчас признано искусственным.
Принимаемое ранее разделение стебельчатобрюхих на паразитических (Parasitica) и жалящих (Aculeata) сейчас признано искусственным.
Несмотря на разноречивость мнений о филогении перепончатокрылых и их положении в системе класса, бесспорно то, что основные этапы их становления были теснейшим образом связаны с покрытосеменными растениями. Их роль как опылителей цветков особенно возросла при появлении в процессе эволюции пчелиных (Apoidea). Столь же определённо мнение о том, что эволюция перепончатокрылых происходила в тропиках, где и сейчас они особенно разнообразны.
Почти все представители отряда размножаются обоеполым путём в сочетании с арренотокным партеногенезом, когда из неоплодотворённых яиц развиваются только самцы. Другие формы партеногенеза встречаются несравненно реже, и лишь у некоторых рогохвостов и пилильщиков наблюдаются телитокия и дейтеротокия. Свойственный перепончатокрылым гаплодиплоидный механизм регуляции
пола потомков, по-видимому, служит предпосылкой формирования семей и общественного образа жизни. Однако для некоторых паразитических перепончатокрылых характерен педогенез, иногда сочетающийся с полиэмбрионией.
Однако для некоторых паразитических перепончатокрылых характерен педогенез, иногда сочетающийся с полиэмбрионией.
В отношении постэмбриональных преобразований все представители отряда характеризуются гистолитическим метаморфозом в фазе куколки, и лишь у некоторых наездников наблюдается гиперметаморфоз. Личинки сидячебрюхих — ложногусеницы — с развитыми грудными ногами и шестью парами брюшных ложноножек — обычно фитофаги, питающиеся либо открыто, либо в плодах и галлах (некоторые Tenthredinidae), либо в стеблях злаков (Cephidae) и древесине (Siricidae). Лишь представители семейства Orussidae паразитируют на насекомых и других членистоногих. Взрослые сидячебрюхие потребляют пыльцу и нектар или вообще не питаются.
Характерные для стебельчатобрюхих червеобразные, лишённые глаз и ног личинки паразитируют на насекомых, или образуют галлы (орехотворки — Cynipoidea), или пожирают свежеубитые и парализованные самкой жертвы. Пчелы и муравьи обычно выкармливают своих личинок предварительно заготовленной пищей. Свойственный им общественный образ жизни характеризуется дифференциацией рабочих особей в зависимости от исполняемых ими функций.
Пчелы и муравьи обычно выкармливают своих личинок предварительно заготовленной пищей. Свойственный им общественный образ жизни характеризуется дифференциацией рабочих особей в зависимости от исполняемых ими функций.
Практическое значение перепончатокрылых в целом положительно, так как многие из них — специфические опылители и естественные регуляторы численности вредных насекомых. Многие виды с успехом используют в практике биологической защиты сельскохозяйственных культур и разводят в промышленных масштабах на биофабриках. Очевидно значение медоносных пчёл как производителей ценнейших пищевых (мёд), лекарственных (пчелиное молочко, прополис, пчелиный яд) и технических (воск) продуктов.
Вредоносность представителей отряда проявляется прежде всего в том, что многие виды пилильщиков и рогохвостов повреждают растения, а некоторые муравьи — строительные материалы и продукты питания. Заметный вред пчеловодству наносят осы-филанты и шершни. Некоторые жалящие перепончатокрылые временами наносят человеку весьма болезненные уколы, чреватые аллергическими реакциями. Перепончатокрылых, в частности пчёл и муравьёв, используют в разноплановых экспериментах.
Перепончатокрылых, в частности пчёл и муравьёв, используют в разноплановых экспериментах.
Перепончатокрылые
1.2. Внешнее строение Голова
Голова, грудь и брюшко резко обособлены кроме сидячебрюхих, у которых брюшко «плотно сидит» на груди. Голова свободная, в основном расширенная в поперечном направлении, гипогнатная ротовые органы направлены вниз или прогнатная … вперёд. Сложные глаза почти всегда хорошо развиты, между ними обыкновенно три расположенных в виде треугольника простых глазка; встречаются также формы, лишённые глазков или совершенно слепые например, рабочие муравьи некоторых видов. На голове выделяют лоб frons, наличник клипеус, темя vertex, затылок occiput, щёки genae, виски tempora.
Усики антенны, или сяжки, развиты в весьма различной степени и имеют от 3 до 60 члеников; они бывают прямые или коленчатые, причём различают стебелёк скапус и прикреплённый к нему под углом жгутик flagellum.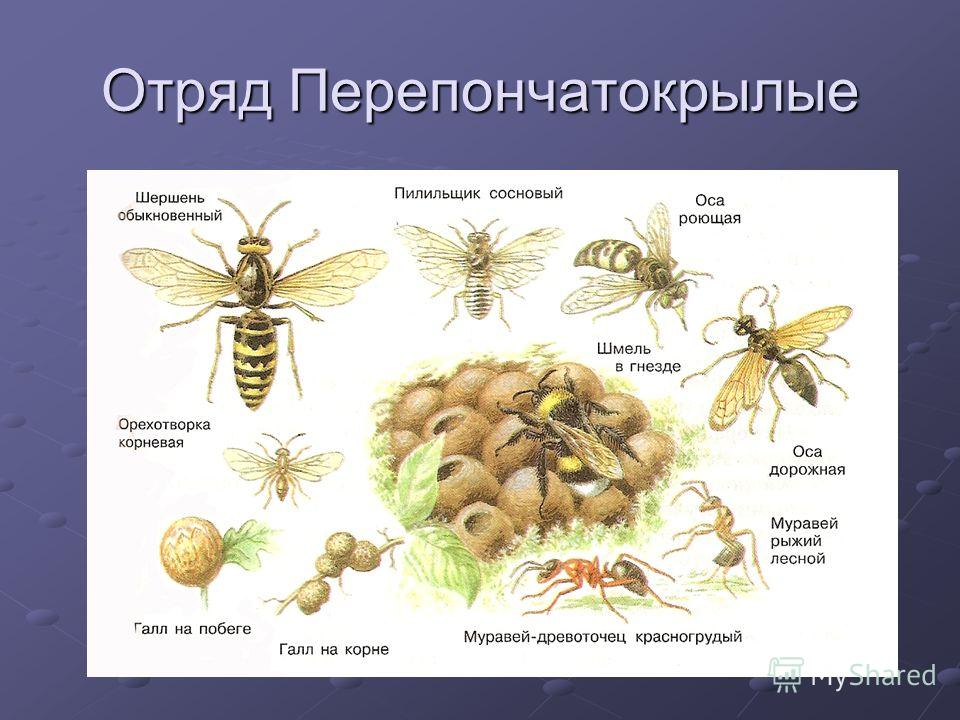 Между ними иногда выделяют поворотный членик, или педицеллюс.
Между ними иногда выделяют поворотный членик, или педицеллюс.
Ротовые органы или чисто грызущего типа, или же нижняя губа labium и нижние челюсти максиллы изменены в органы принятия жидкой пищи органы лизания, лакания, образуя единый лабио-максиллярный комплекс. Сильно развитые верхние челюсти мандибулы, или жвалы у некоторых служат для измельчения пищи, а у большинства — орудиями при постройке жилищ, прогрызании ходов в дереве прогрызали и свинцовые листы, приготовления пищи для личинок, убивания добычи, таскания строительных и питательных материалов и т. п. На нижних челюстях и нижней губе расположены нижнечелюстные максиллярные щупики 6-члениковые или менее и нижнегубные лабиальные щупики 4-члениковые или менее.
Мозг рабочих пчёл Apis содержит около 850 000 нейронов и занимает 0.35 % — 1.02 % от общего объёма тела. У муравьёв рода Formica мозг занимает 0.57 % объёма тела. У одного из самых маленьких наездников Megaphragma mymaripenne его длина 200 мкм в мозгу только 4600 нейронов, из которых лишь 5 % содержат хромосомы.
Изменения в фауне перепончатокрылых насекомых Ростовской области и прогноз экологических последствий
Инвазийные виды считаются второй по значимости угрозой биоразнообразию [14]. Проникновение новых инвазийных организмов происходит постоянно и независимо от их карантинного статуса и на территории России в последние годы приобретает характер национальной проблемы. Объектами их воздействия становятся природные биоценозы и агроэкосистемы.
Перепончатокрылые насекомые имеют огромное влияние на экосистемы как фитофаги (многочисленные виды пилильщиков), энтомофаги (представители муравьев) и опылители (в основном дикие одиночные пчелы)
В Ростовской области в 2011 году наблюдались сильные повреждения вяза личинками неизвестного в европейской части России пилильщика, который позднее был идентифицирован как Aproceros leucopoda (Takеuchi, 1939)-сем. Argidae — восточноазиатский ильмовый пилильщик-зигзаг, пилильщик бледноногий [an East Asian sawfly].
Естественный ареал ильмового пилильщика охватывает Японию, восток Китая, Корейский полуостров и российский Дальний Восток [1]. В начале ХХI века этот вид был выявлен в Китае, более чем в тысяче км от его естественного ареала [15]. В 2003 году этот фитофаг был впервые отмечен в Венгрии и Польше [2]. В 2009 году он уже широко распространился в Венгрии [4, 7], в 2005 году был выявлен в Румынии, в 2006 году — в восточной части Луганской области (Украина) вблизи российской границы. В настоящее время он проник также в Австрию, Словакию [2]
В начале ХХI века этот вид был выявлен в Китае, более чем в тысяче км от его естественного ареала [15]. В 2003 году этот фитофаг был впервые отмечен в Венгрии и Польше [2]. В 2009 году он уже широко распространился в Венгрии [4, 7], в 2005 году был выявлен в Румынии, в 2006 году — в восточной части Луганской области (Украина) вблизи российской границы. В настоящее время он проник также в Австрию, Словакию [2]
В европейской части России этот вид впервые выявлен в посадках вяза мелколистного (Ulmus pumila L.) Краснодарского края. В 2010 году на Кубани локальное массовое размножение пилильщика фиксировалось преимущественно в полезащитных лесополосах и искусственных насаждениях вдоль федеральных шоссе [5].
Нами в 2011 году во многих районах Ростовской области отмечена широкомасштабная вспышка численности пилильщика. В июле была отмечена сильная дефолиация вяза на протяжении сотен километров полезащитных и придорожных насаждений от границы с Краснодарским краем и почти до границы с Воронежской областью. Площадь поврежденных насаждений менее чем за месяц увеличилась в несколько раз. Максимальная плотность личинок и площадь очагов отмечены как в южных, так и северо-западных районах области. Вся западная половина Ростовской области оказалась заселена ильмовым пилильщиком. Очень высокая численность насекомого привела к полной потере листьев вяза в середине лета. В Ростовской области наблюдалось состояние вязовых насаждений с частичным усыханием крон. Территория, на которой в 2011 году отмечалось присутствие пилильщика в насаждениях охватывает почти 30 тысяч км2.
Площадь поврежденных насаждений менее чем за месяц увеличилась в несколько раз. Максимальная плотность личинок и площадь очагов отмечены как в южных, так и северо-западных районах области. Вся западная половина Ростовской области оказалась заселена ильмовым пилильщиком. Очень высокая численность насекомого привела к полной потере листьев вяза в середине лета. В Ростовской области наблюдалось состояние вязовых насаждений с частичным усыханием крон. Территория, на которой в 2011 году отмечалось присутствие пилильщика в насаждениях охватывает почти 30 тысяч км2.
Обширная площадь инвазии этого фитофага в Ростовской области не оставляет сомнения в его расселительных способностях, что делает восточноазиатского ильмового пилильщика опасным вредителем вяза в искусственных насаждениях Ростовской области и Юга России. Вероятно, вид проник в регион несколько лет назад. Учитывая то, что пилильщик заселил только западную половину области можно предположить, что инвазия происходила несколькими путями со стороны Приазовья и Украины. Наиболее активное расселение вредителя наблюдается вдоль автомагистралей, окруженных посадками вяза, к которым примыкают полезащитные лесополосы с участием вяза, представляющие коридоры для расселения вида по степной зоне.
Наиболее активное расселение вредителя наблюдается вдоль автомагистралей, окруженных посадками вяза, к которым примыкают полезащитные лесополосы с участием вяза, представляющие коридоры для расселения вида по степной зоне.
Особенности биологии этого фитофага на юге России исследованы недостаточно полно. Лёт имаго начинается обычно в начале мая (в 2012 в конце апреля), позже (до конца сентября) в древостоях можно встретить все стадии развития пилильщика. В сентябре — октябре 2011 года активные фазы пилильщика в его очагах на территории области не были обнаружены, несмотря на высокую численность вида в июле и начале августа.
Пилильщики развиваются в трех генерациях уже к середине июля на юге и в центральных районах Ростовской области. Самки откладывают яйца в вершинки зубчиков листьев, личинки активно питаются в течение двух недель. Личинки наносят повреждения в виде характерных «зигзагообразных» погрызов листовой пластинки. При высокой плотности они съедают молодой лист до основания черешка.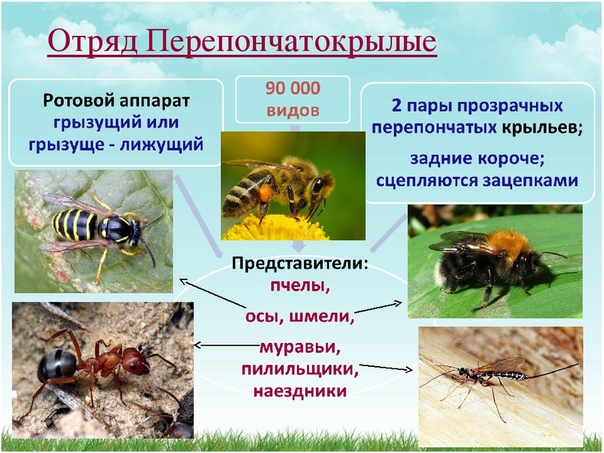 Летние поколения окукливаются на листьях, ветвях, и в трещинах коры на стволах в рыхлом коконе. Фаза куколки второй генерации продолжается 4–5 суток. Выход имаго второй генерации завершается за 4–5 дней.
Летние поколения окукливаются на листьях, ветвях, и в трещинах коры на стволах в рыхлом коконе. Фаза куколки второй генерации продолжается 4–5 суток. Выход имаго второй генерации завершается за 4–5 дней.
Осенью коконы локализуются в подстилке. Зимует пилильщик в коконах, которые личинки, уходящие на зимовку, формируют в подстилке и верхних слоях почвы. Такие коконы имеют светло-коричневый цвет, более плотное строение и инкрустированы частичками почвы. Численность пилильщика от поколения к поколению возрастает. Дефолиация насаждений к концу питания первой генерации хорошо различима при плотности коконов 20–30 на 100 ростовых точек. К моменту окукливания личинок второй генерации при плотности коконов (обеих генераций) в 100–130 на 100 ростовых точек она достигает сплошной степени [5]. Однако повреждения наносят не коконы, а личинки и поэтому вредоносность необходимо оценивать именно для них. Коконы удобны для учетов и могут использоваться для прогнозирования популяции вредителя. В наших условиях 1–2 личинки на лист уже представляли серьезную угрозу деревьям. Реальная численность личинок превышала и 10 экземпляров на один лист.
В наших условиях 1–2 личинки на лист уже представляли серьезную угрозу деревьям. Реальная численность личинок превышала и 10 экземпляров на один лист.
Постоянные повреждения пилильщиком могут привести к исчезновению видов вяза из лесных насаждений региона.
Муравьи, как консументы, в экосистемах выполняют очень разнообразные функции. Роль разных видов варьирует от зоофагии до фитофагии с различными переходными вариантами.
Отмечен новый вид муравья Lasius neglectus (Hymenoptera: Formicidae) этот Вид муравья описан в работе [13], на территории Венгрии. С тех пор сообщения о его находках поступили из многих стран Восточной, Южной и Центральной Европы, Турции и Грузии [10], из Великобритании [12], из Турменистана, Узбекистана [11]. Работа [13] содержит упоминание о находке Lasius neglectus на территории черноморского побережья России в 1984 году (точка расположена вблизи Сочи и указана как территория Грузии, вероятно, ошибочно).
Вид продолжает расширение своего ареала и пагубно влияет на биоразнообразие, в местах, где его колонии доминируют, численность прочих муравьев, видов собирающих пищу на поверхности земли (стратобионтов) и растительности, сокращается. На численность других видов членистоногих (в том числе сожителей и тлей) L. neglectus также может оказывать влияние [9].
На численность других видов членистоногих (в том числе сожителей и тлей) L. neglectus также может оказывать влияние [9].
Как минимум в некоторых колониях вид может проникать в жилища и хозяйственные постройки, фуражируя в них (в частности, два раза был замечен в теплицах), но предпочитает поселение снаружи, чаще встречаясь в садах и на приусадебных хозяйствах, где может провоцировать вспышки численности равнокрылых (обычно тлей).
Образцы собирались непосредственно из гнезд или же с посещаемых для сбора пади растений. Определение осуществлялось по описанию вида в [13], изучение и измерение размеров тела — при помощи бинокуляра с измерительной сеткой.
Гнезда вида были обнаружены в трех точках в границах г. Ростова-на-Дону (Зеленый остров, Ботанический сад, территория Набережной) и в нескольких близко расположенных точках на территории г. Батайск. Всего обнаружено полигинных гнезд шесть. Гнезда чаще располагаются под камнями, строительным мусором, стволами деревьев, частично могут располагаться под корой деревьев и в земле.
Сбор по крупнейшей, из известных автору, колонии осуществлялся в течение трех лет (2009–2012) с марта и до окончания активности (октябрь -ноябрь). Остальные колонии были найдены в теплое время 2012 года.
Вид во многом похож на широко распространенный в России вид Lasius alienus Forster, 1850 и достоверно может быть отличен по выраженной полигинности (наличию в одной колонии множества плодущих самок, а не одной как при моногинности), не характерной для других европейских видов Lasius. Помимо полигинности представители колоний отличаются размером самцов: средняя длина тела для Lasius neglectus = 2.9 мм, наибольшая = 3.6 мм, у взятых для сравнения самцов L. alienus все размеры больше: средняя длина тела = 4.3 мм, наибольшая = 4.8 мм.
Рабочие особи внешне сходны, отличаются у L. neglectus менее выраженным килем наличника (чаще не выраженным вообще) и числом зубцов мандибул: для L. neglectus обычное число зубцов — 7, из которых основных — обычно 2, редко 3; для сравниваемых особей L. alienus обычное число зубцов — 9, из которых основных обычно — 3. Цвет рабочих L. neglectus несколько более светлый, чем у L. alienus, размеры — незначительно меньше.
alienus обычное число зубцов — 9, из которых основных обычно — 3. Цвет рабочих L. neglectus несколько более светлый, чем у L. alienus, размеры — незначительно меньше.
Размер самок также меньше чем у других Lasius и в среднем составляет 5.8 мм (у взятых для сравнения L. alienus средний размер = 7.4 мм).
Рис. 1 — Королевы Lasius neglectus в окружении рабочих (вид снизу) и рабочие крупным планом
В наибольшем из наблюдаемых гнезд, найденном под листами рубероида, максимально регистрировалось до 30–40 плодущих самок одновременно, хотя их общее число может быть значительно большим (порядка десятков тысяч) [8], так как крупное гнездо продолжается в земле и дерновинах травы. В меньших гнездах обнаруживались порядка 10 плодущих самок. В осматриваемом гнезде наблюдалось значительное число крылатых самок и самцов с 02.06.12 по 15.06.12, и с 31.05.11 по 11.06.11, в том числе — спаривающихся. При вскрытии гнезда половые особи демонстрировали бегство в более глубоко расположенные части гнезда. Брачный лёт не наблюдался, к концу периода наблюдения в гнезде и его окрестностях находилось большое число бескрылых самок и мертвых самцов.
Брачный лёт не наблюдался, к концу периода наблюдения в гнезде и его окрестностях находилось большое число бескрылых самок и мертвых самцов.
Окружение найденных гнезд представляет собой умеренно увлажненные стации с близко расположенной древесно-кустарниковой растительностью (лесополосы, посадки ботанического сада, одичавшая кустарниковая растительность — тополь, тёрн, алыча и др.). Рабочие особи активно участвуют в сборе сахаристых веществ: пади тлей и иногда нектара цветов на таких растениях как чертополох, бодяк и осот.
Две колонии располагались в садах, непосредственно примыкающих к человеческим жилищам и хозяйственным постройкам, в этих случаях отдельные рабочие особи обнаруживались в домах и хозяйственных постройках.
Обычно такие виды муравьев как Formica sanguinea, Formica cunicularia, Lasius niger, Camponotus piceus встречались на той же территории, что и гнезда данного вида. В пределах одного гнезда наблюдалась колония Solenopsis fugax. Рядом с гнездами, обнаруженными в городских парках, идентифицировались, главным образом, представители Lasius niger, L. alienus, L. brunneus, Crematogaster subdentata.
alienus, L. brunneus, Crematogaster subdentata.
В гнездах были зарегистрированы такие сожители, как Myrmicophila, личинки кожеедов, многочисленные особи щетинохвосток (Atelura sp.). Большое количество плодущих самок и рабочих Lasius neglectus заражено клещами. Сожители очень многочисленны, по сравнению с их количеством в гнездах других Lasius, их высокая численность и распространенность в гнездах может быть связана с расселением L. neglectus без брачного лёта и большей, чем у других видов Lasius, общей численностью муравьев в гнезде.
Можно предположить, что вскоре вид будет обнаружен в большинстве крупных городов и южных регионов России. Дальнейшее распространение вида может привести к обеднению видового разнообразия муравьев, нарушению сложившихся с ними экологических связей других видов, причинению некоторого ущерба садовым, тепличным культурам. Большая численность и несколько меньшие, чем у местных видов, рабочие особи, возможно приведут к увеличению поражений растений мирмекофильными равнокрылыми.
Миллионы лет коэволюция цветковых растений зависела и определялась взаимодействием в системе растение — пчела-опылитель.
Одиночные пчелиные (Hymenoptera: Apoidea) являются основными опылителями главных энтомофильных культур в нашей стране — подсолнечника и люцерны. В Ростовской области более 500 видов пчелиных и среди них около 100 экономически значимых опылителей [6]. Численность опылителей в настоящее время оценивается как достаточная для полного опыления растений. Период лёта экономически значимых видов опылителей охватывает время с мая по август. Продолжительность цветения культур короче периода лёта опылителей. Пчелиные концентрируются на энтомофильных культурах только в фазу их цветения и с учетом пищевой специализации: например в Ростовской области часть видов пчел концентрируется в мае на эспарцете, в июне на люцерне, в июле — на подсолнечнике.
Последнее время для борьбы с вредителями регистрируются высокотоксичные для пчел инсектициды из группы неоникотиноидов [3]. В экологических ограничениях по применению этих препаратов есть регламенты по температуре и пространственной изоляции в 5 км для медоносной пчелы (Apis mellifera, Linnaeus 1758), но одиночные пчелы в них никак не учитываются.
В экологических ограничениях по применению этих препаратов есть регламенты по температуре и пространственной изоляции в 5 км для медоносной пчелы (Apis mellifera, Linnaeus 1758), но одиночные пчелы в них никак не учитываются.
Нами выявлены факты отрицательного влияния применения неоникотиноидов на опылителей и урожай энтомофильных культур. Обработка посевов пшеницы против личинок вредной черепашки на территории одного хозяйства приводит к гибели одиночных пчел и отсутствию опылителей даже в соседних хозяйствах и снижению урожая подсолнечника и люцерны. Урожайность семян люцерны снизилась до 30 кг с га, что ниже обычного почти в 10 раз.
Использование экотехнологий позволяет уменьшить пестицидные нагрузки и сохранить опылителей и энтомофагов при проведении защитных мероприятий. Но при продолжении практики игнорирования экологических ограничений в применении пестицидов прогнозируется тяжелая ситуация для опылителей и снижение урожаев энтомофильных культур и уменьшение биоразнообразия растений и насекомых.
Заключение
Естественные и антропогенные изменения в фауне перепончатокрылых могут привести к кардинальному изменению облика региона, сильному изменению структуры агроэкосистем и снижению урожайности энтомофильных сельскохозяйственных культур.
Необходимо включить упомянутые новые виды в список фауны Ростовской области и юга России, определители по имаго и личинкам. Продолжить исследования по их биологии, по уточнению ареалов и изучению их естественных врагов на юге России.
Мы уже познаем структуру и закономерности функционирования эко- и агроэкосистем. Методы прогнозирования экологических проблем дают возможность предвидеть последствия деятельности человека даже в энтомологических аспектах, что является движением к управлению ноосферой, о чем мечтал Владимир Иванович Вернадский.
Список литературных источников
- Артохин К. С. Энтомоценоз люцерны. Ростов на Дону, 2000. – 204с.
- Масляков В. Ю., Ижевский С.
 С. Инвазии растительноядных насекомых в европейскую часть России. М. : ИГРАН, 2011. – 272 с.
С. Инвазии растительноядных насекомых в европейскую часть России. М. : ИГРАН, 2011. – 272 с. - Список пестицидов и агрохимикатов, разрешенных к применению на территории Российской Федерации. М., 2011. – 953 с.
- Сундуков Ю. И. Подотряд Symphyta — сидячебрюхие. Насекомые Лазовского заповедника. Владивосток: Дальнаука, 2009. — С. 212—220.
- Щуров В. И., Гниненко Ю. И., Ленгесова Н. А., Гниненко М. Ю. Ильмовый пилильщик в европейской России. //Защита и карантин растений. 2012, №2. — С. 37—38.
- Blank S. M., Hara H., Mikulas J., Csoka G., Ciornei C., Constantineanu R., Constantineanu I., Roller L., Altenhofer E., Huflejt T., Vetek G. Aproceros leucopoda (Hymenoptera: Argidae): An East Asian pest of elms (Ulmus spp.) invading Europe //Eur. J. Entomol., 2010. 107. – Р. 357—367.
- Csoka G. Mikulas J., Blank S., Vetek G. A kanjargos szilleveldarazs (Aproceros leucopoda Takeuchi, 1939) megjelenese Magyarorszagon. //Novenjvedelmi Tudomangos Napok 2010, Budapest, 2010.
 — P. 19—20.
— P. 19—20. - Espadaler X.; Rey S.; V.Bernal. 2004. Quenn number in supercolony of the invasive garden ant Lasius neglectus Insectes sociaux 51 (3). — 232 p.
- Fox, M. 2010. First incursion of Lasius neglectus (Hymenoptera: Formicidae), an invasive polygynous ant in Britain. British Journal of Entomology and Natural History 23: 1803. — Р. 1—3.
- Paris, C. & Espadaler, X. 2012. Foraging activity of native ants on trees in forest fragments colonized by the invasive ant Lasius neglectus. Psyche 2012 (261316). — Р. 1—9.
- Schultz, R. & B. Seifert. 2005. Lasius neglectus (Hymenoptera: Formicidae) — a widely distributed tramp species in Central Asia. Myrmecologische Nachrichten 7. — Р. 47—50.
- Seifert, B. 2000. Rapid range expansion in Lasius neglectus (Hymenoptera, Formicidae) — an Asian invader swamps Europe. Dtsch. Entomol. Z., 47. — Р. 173—179.
- Van Loon, A. J.; Boomsma, J. J.; Andrasfalvy, A. 1990. A new polygynous Lasius species (Hymenoptera: Formicidae) from central Europe.
 I. Description and general biology. Insectes Soc. 37: pp. 348—362 (page 350, fig. 5 worker, queen, male described)
I. Description and general biology. Insectes Soc. 37: pp. 348—362 (page 350, fig. 5 worker, queen, male described) - Vetek G., Mikulas J., Csoka G., Blank S. A kanjargos szilleveldarazs (Aproceros leucopoda Takeuchi, 1939) magyarorszagon. //Novenyvedelem, 2010, v. 46, N11. — Р. 519—520.
- Wu Xing-yu, Xin Heng A new record of the genus Aproceros Malaise (Hymenoptera: Argidae) from China //Entomotaxonomia, 2006, v. 28, №4. — Р. 279—280.
Библиографическая ссылка
Артохин К. С., Игнатова П. К., Колесников С. И., Решетов А. A. Изменения в фауне перепончатокрылых насекомых Ростовской области и прогноз экологических последствий // «Живые и биокосные системы». – 2013. – №2; URL: http://www.jbks.ru/archive/issue-2/article-6.
Mutillidae, Vespidae, Apidae) в питании азиатского барсука (meles leucurus Hodgson, 1847) на Урале и в Западной Сибири – тема научной статьи по биологическим наукам читайте бесплатно текст научно-исследовательской работы в электронной библиотеке КиберЛенинка
333^^ Аграрный вестник Урала №6 (85), 2011 г. +XSSx.
+XSSx.
Биология
жалящие перепончатокрылые
(HYMENOPTERA: MUTILLIDAE, VESPIDAE, APIDAE) В ПИТАНИИ АЗИАТСКОгО БАРСУКА (MELES LEUCURÜS HODGSON, 1847;
на урале и в западной сибири
о. С. ЗАГАйновА, аспирант,
п. в. рудоискатель,
научный сотрудник, институт экологии растений и животных Уро РАИ
620144, г. Екатеринбург, ул. 8 Марта, д. 202; тел. 8 (343) 210-38-55 доб. 140; e-mail: [email protected]
Ключевые слова: барсук, питание, осы, шмели, Meles leucurus, Hymenoptera, Vespidae, Mutillidae, Apidae. Keywords: badger, diet, wasps, bumblebees, Hymenoptera, Vespidae, Mutillidae, Apidae.
Межвидовые отношения в биоценозах реализуются через сложные формы взаимодействия популяций разных видов, в основе которых лежат трофические связи [13]. Необходимость детального изучения питания и трофоценотических связей животных определяется не только первостепенной биологической ролью этой стороны их жизнедеятельности, но и влиянием, которое они оказывают на многие стороны экологии и этологии животных [9].
Барсук — всеядный хищник. Анализ географической динамики структуры его рациона представляет особый интерес, поскольку эти изменения отражают изменения как трофических потребностей и предпочтений самого потребителя, так и состояния потенциальных кормовых объектов [7]. В большинстве работ, посвященных изучению его питания, приводятся сведения об общем спектре кормов. В то же время, оценка роли конкретной таксономической группы жертв в рационе вида позволит более точно охарактеризовать его роль в различных природных сообществах. leucurus Hodgson,1847) констатируется только факт присутствия ос и шмелей в его рационе, тогда как сведения об их видовом составе и встречаемости не приводятся [3, 8, 12].
leucurus Hodgson,1847) констатируется только факт присутствия ос и шмелей в его рационе, тогда как сведения об их видовом составе и встречаемости не приводятся [3, 8, 12].
Целью работы было охарактеризовать трофическое звено «барсук-жалящие перепончатокрылые» на Урале и в Западной Сибири. В частности, выяснить видовой состав жертв, отмеченных в питании хищника в районах исследований, провести сравнительный анализ географической изменчивости их встречаемости и обилия. Также была предпринята попытка на основании данных о таксономическом составе и биологии перепончатокрылых описать особенности добывания их барсуком.
Методика исследований.
Сбор материала (экскременты барсука) проводили: 1) в 1999-2005 гг. в конце июня-начале июля в северной лесостепи Среднего Урала (Свердловская область, Каменский район, окрестности дер. Старикова) — 151 проба; 2) в конце июня-начале июля в 2004, 2009, 2010 гг. в южной тайге Зауралья (Свердловская область, Талицкий район, национальный парк «Припышминские боры») — 77 проб;
в южной тайге Зауралья (Свердловская область, Талицкий район, национальный парк «Припышминские боры») — 77 проб;
3) в 2004-2010 гг. в конце мая-начале июня («весна») и в начале августа-в сентябре («осень») в северной лесостепи в средней тайге Западной Сибири (Ханты-Мансийский АО, природный парк «Самаровский чугас», острова Большой и Малый Чухтинские) — 255 проб.
Пробы промывали водой через стандартную колонку почвенных сит, далее просушивали на солнце. Камеральная обработка заключалась в разборе непереваренных остатков. По сохранившимся хитиновым покровам определяли таксономическую принадлежность и подсчитывали количество особей жертв. Определение проводилось большей частью по головным капсулам насекомых, имеющим наилучшую сохранность.
Для каждого района исследований оценивали общую встречаемость кормовых объектов (процент проб с данным видом корма от общего числа проб за весь период наблюдений). ае) и не менее 6 видов семейства складчатокрылых (бумажных) ос (Vespidae) (табл. 1). Все осы были представлены исключительно самками. Пчелиные (Apidae) были определены лишь до семейства, скорей всего, барсук питался шмелями (род Bombus Latreille, 1802), но возможно, он потреблял шмелей-кукушек (род Psithyrus Lepeletier, 1833). У этих двух родов головные капсулы имеют сходное строение,
ае) и не менее 6 видов семейства складчатокрылых (бумажных) ос (Vespidae) (табл. 1). Все осы были представлены исключительно самками. Пчелиные (Apidae) были определены лишь до семейства, скорей всего, барсук питался шмелями (род Bombus Latreille, 1802), но возможно, он потреблял шмелей-кукушек (род Psithyrus Lepeletier, 1833). У этих двух родов головные капсулы имеют сходное строение,
Таблица 1
Таксономический состав перепончатокрылых в пробах
Район
Таксон Средний Урал Зауралье Западная Сибирь
Семейство Mutillidae Осы — немки
Mutilla europaea Linnaeus, 1758 + — —
Семейство Vespidae Складчатокрылые осы
Vespa crabro Linnaeus, 1758 + + +
Dolichovespula saxonica (Fabricius, 1793) + +
Dolichovespula norvegica (Fabricius, 1781) — — +
Vespula vulgaris (Linnaeus, 1758) + + +
Vespula rufa (Linnaeus, 1758) + + +
Vespula germanica (Linnaeus, 1793) — — +
Семейство Apidae Пчелиные
Bombus Latreille, 1802 или Psithyrus Lepeletier, 1833 | 1 + 1 1 + 1 1 +
Примечание: «+» или «-
— присутствие или отсутствие вида соответственно Таблица 2
Обилие особей перепончатокрылых в пробах
Район Показатель
число проб число особей
общее в выборке среднее на пробу ± ошибка максимальное на пробы
Семейство Vespidae
Средний Урал 15 28 1,9 ± 1,46 6
Зауралье 9 14 1,8 ± 0,28 3
Западная Сибирь «весна» 12 12 1,1 ± 0,08 2
Западная Сибирь «осень» 10 20 2,0 ± 0,68 7
Семейство Apidae
Средний Урал 55 121 2,2 ± 2,25 12
Зауралье 28 70 2,5 ± 0,51 11
Западная Сибирь «весна» 25 31 1,3 ± 0,13 3
Западная Сибирь «осень» 33 35 1,1 ± 0,04 2
»
¡55^*^ Аграрный вестник Урала №6 (85), 2011 г. Па europaea Ь. отмечено только на Среднем Урале — в одной пробе 6 особей, в другой 1 особь. В большинстве случаев барсук использовал в питании от 1 до 7 особей Vespidae, от 1 до 12 особей Apidae. В то же время были пробы с большим числом представителей Vespidae. Так, на Среднем Урале в одной пробе обнаружено 15 особей, в Зауралье — 51, в Западной Сибири в осенний период в одной — 21, а в другой — 46 особей. В табл. 2 приведены показатели обилия этих семейств, среднее и максимальное число особей в пробе даны без учета выше обозначенных случаев массового потребления.
Па europaea Ь. отмечено только на Среднем Урале — в одной пробе 6 особей, в другой 1 особь. В большинстве случаев барсук использовал в питании от 1 до 7 особей Vespidae, от 1 до 12 особей Apidae. В то же время были пробы с большим числом представителей Vespidae. Так, на Среднем Урале в одной пробе обнаружено 15 особей, в Зауралье — 51, в Западной Сибири в осенний период в одной — 21, а в другой — 46 особей. В табл. 2 приведены показатели обилия этих семейств, среднее и максимальное число особей в пробе даны без учета выше обозначенных случаев массового потребления.
На Среднем Урале и Зауралье отмечены сходные значения по среднему числу особей на пробу для Vespidae и Apidae. Для Урала этот показатель по двум семействам выше, чем для Западной Сибири. Среднее число особей на пробу Vespidae в Западной Сибири в весенний период выше, чем в осенний, тогда как для Apidae отмечена противоположная тенденция.
Обсуждение.
По нашим данным, на Урале и в Западной Сибири выявлена относительно высокая встречаемость жалящих перепончатокрылых в питании азиатского барсука (в среднем около 37,8 %), при этом процент встреч Apidae (в среднем около 30,5 %) выше, чем у Vespidae (в среднем около
13.9 %). В рационе европейского барсука общая встречаемость жалящих перепончатокрылых несколько ниже, главную роль играют осы. В Швейцарии встречаемость шмелей составляла 1 %, ос — 13 % [16], в Молдавии процент встреч шмелей — 0,5 %, ос — 2,2 % [4], в Крыму процент встреч шмелей — 6 %, ос — 18 % [1].
Роль перепончатокрылых в питании барсука может меняться в зависимости от сезона. По нашим данным, в Западной Сибири наблюдается небольшой рост потребления Apidae в осенний период, по сравнению с весенним, тогда как для Vespidae отмечена противоположная тенденция (рис.1). зо пончатокрылых, отмеченное в районах | наших исследований, а также в других | 2о частях ареала, указывает на то, что бар- | сук, вероятно, целенаправленно отыски- I 10 вает и раскапывает гнезда шмелей и ос. К середине лета в гнездах содержатся сотни о личинок и куколок, повышающих их привлекательность. Нами было обнаружено, что в южной тайге Среднего Урала гнезда ос (Vespa crabro L., Dolichovespula saxonica F., Vespula vulgaris L.) к началу сентября могут насчитывать до 1300 особей (включая рабочих, цариц следующего года и самцов), свыше тысячи личинок и нескольких сот куколок. Масса живого вещества таких гнезд может достигать 1 кг
зо пончатокрылых, отмеченное в районах | наших исследований, а также в других | 2о частях ареала, указывает на то, что бар- | сук, вероятно, целенаправленно отыски- I 10 вает и раскапывает гнезда шмелей и ос. К середине лета в гнездах содержатся сотни о личинок и куколок, повышающих их привлекательность. Нами было обнаружено, что в южной тайге Среднего Урала гнезда ос (Vespa crabro L., Dolichovespula saxonica F., Vespula vulgaris L.) к началу сентября могут насчитывать до 1300 особей (включая рабочих, цариц следующего года и самцов), свыше тысячи личинок и нескольких сот куколок. Масса живого вещества таких гнезд может достигать 1 кг
Как уже отмечалось ранее, сведения о видовом составе жалящих перепончатокрылых в питании барсука весьма фрагментарны. Так, в Германии выявлены следующие виды ос: Vespula vulgaris L., V. rufa L., Vespa crabro L., Psithyrus Lep. и Anthophora Latr. [14]. Первые три вида также отмечены в питании барсука в наших районах исследований. Потребление барсуком шмелей Bombus lucorum L. (анализ содержимого желудка) показано в Англии [18]. Любопытные сведения по анализу содержимого желудка барсуков, добытых в широколиственных лесах Тульских засек, приводит Г. Л. Лихачев [6]. У одного зверя в желудке была 51 взрослая оса и 42 ее личинки (Vespa vulgaris L.), кроме того, сохранилось несколько кусочков шмелиных сот, не менее 50 ячеек (Bombus terrestis L.), у другого было обнаружено 362 ячейки сот с молодыми, готовыми к вылету особями и некоторое количество взрослых шмелей, а кроме того, 15 ос.
Потребление барсуком шмелей Bombus lucorum L. (анализ содержимого желудка) показано в Англии [18]. Любопытные сведения по анализу содержимого желудка барсуков, добытых в широколиственных лесах Тульских засек, приводит Г. Л. Лихачев [6]. У одного зверя в желудке была 51 взрослая оса и 42 ее личинки (Vespa vulgaris L.), кроме того, сохранилось несколько кусочков шмелиных сот, не менее 50 ячеек (Bombus terrestis L.), у другого было обнаружено 362 ячейки сот с молодыми, готовыми к вылету особями и некоторое количество взрослых шмелей, а кроме того, 15 ос.
Наличие исключительно самок жалоносных перепончатокрылых в наших районах исследований в июньских и июльских пробах объясняется тем, что первые самцы шмелей и бумажных ос появляются на Среднем Урале к середине II — началу III декады июля. Среди Vespidae, по-видимому, только один вид (Vespula rufa L.) имеет укороченный цикл развития, для которого отмечено раннее появление самцов (конец июня-начало июля) и завершение лета этого вида уже к началу сентября. Кроме того, наличие самок ос и шмелей в питании барсука говорит о том, что он, скорей всего, безразличен к их ужалениям.
Кроме того, наличие самок ос и шмелей в питании барсука говорит о том, что он, скорей всего, безразличен к их ужалениям.
Исходя из особенностей биологии жертв, можно сделать ряд предположений о способах добывания барсуком разных видов жалящих перепончатокрылых. Во-первых, он может раскапывать их гнезда. Известно, что осы рода Vespula Thomson, 1869 живут в земле, занимая, как правило, различные ниши, пустоты, трухлявые пни или норы грызунов. На Урале нами неоднократно отмечены случаи гнездования под землей Vespa crabro L. [10]
1
JA
■ Mutillidae
□ Vespidae
□ Apidae
Д
С ред ний У рал Зураль е
Западная
Сибирь
«весна»
Западная район Сибирь «осень»
Рисунок 1
Встречаемость перепончатокрылых в пробах (за весь период исследований) и Dolichovespula saxonica F. Во-вторых, барсук может питаться осами непосредственно с субстрата. Некоторые виды ос родов Vespa L. и Dolichovespula Rohwer строят гнезда, как правило, над землей. Возможно, барсук разорял упавшее гнездо с этими осами, сбитое другими животными, или находил их в поваленном дереве, в котором они заселились. Самки немок Mutilla europaea L. бескрылые, на Урале этот вид довольно редок. Возможно, барсук потреблял их с земли или, что более вероятно, разорил гнездо шмелей, где они могли находиться в качестве паразитов [5].
Во-вторых, барсук может питаться осами непосредственно с субстрата. Некоторые виды ос родов Vespa L. и Dolichovespula Rohwer строят гнезда, как правило, над землей. Возможно, барсук разорял упавшее гнездо с этими осами, сбитое другими животными, или находил их в поваленном дереве, в котором они заселились. Самки немок Mutilla europaea L. бескрылые, на Урале этот вид довольно редок. Возможно, барсук потреблял их с земли или, что более вероятно, разорил гнездо шмелей, где они могли находиться в качестве паразитов [5].
С. В. Кириковым [3] подчеркивается положительная роль барсука в истреблении общественных ос на том основании, что Vespidae уничтожают множество пчел и других насекомых-опылителей растений. Нам это представляется не совсем обоснованным, поскольку, во-первых, осы сами являются опылителями, во-вторых, осы истребляют насекомых, многие из которых — вредители сельского и лесного хозяйств или имеют отрицательное ветеринарное и медицинское значение.
Обобщая полученные результаты, можно заключить, что жалящие перепончатокрылые играют важную роль в питании барсука на Урале и в Западной Сибири. Их массовое потребление в значительной степени связано с общественным образом жизни жертв и, как следствие, высокой доступностью и локальным нахождением в определенном месте. Хищник поедает сразу большое количество особей и тем самым снижает свои энергетические затраты на кормодобывание. Несмотря на относительно разнообразный состав Vespidae в рационе барсука, скорей всего, он не отдает предпочтения каким-то определенным видам.
Авторы выражают свою благодарность Н. И. Маркову, Е. В. Бердышевой, А. В. Нестеркову за помощь в сборе материала и Е. В. Зиновьеву, определившему часть сборов.
Работа выполнена при под -держке программы развития ведущих научных школ (НШ-3260.2010.4) и научнообразовательных центров (контракт 02. 740.11.0279).
740.11.0279).
литература
1. Альгульян С. Г Питание и сезонные особенности образа жизни крымского барсука (Meles meles Tauricus Одп.) // Зоологический журнал. 1940. Т. 19. Вып. 3. С. 499-515.
2. Данилов П. И., Туманов И. Л. Куньи Северо-Запада СССР. Л., 1976. 256 с.
3. Кириков С. В. Птицы и млекопитающие в условиях ландшафтов южной оконечности Урала. М. : Академия наук СССР, 1952. 411 с.
4. Корчмарь Н. Д. Некоторые данные о распространении, биологии и хозяйственном значении барсука в Молдавии // Вопросы экологии и практического значения птиц и млекопитающих Молдавии. Кишинев, 1962. С. 56-63.
5. Лелей А. С. Осы-немки (Hymenoptera, Mutillidae) фауны СССР и сопредельных стран. Л. : Наука, 1985. 268 с.
Л. : Наука, 1985. 268 с.
Аграрный вестник Урала №6 (85), 2011
___________________Ветеринария ‘У
6. Лихачев Г. Л. Некоторые черты экологии барсука в широколиственных лесах Тульских засек (Приокско-Террасный заповедник) // Сборник материалов по результатам изучения млекопитающих в государственном заповеднике. Москва, 1956. С. 72-78.
7. Марков Н. И., Загайнова О. С., Зиновьев E. B. Предварительные данные о питании барсука в природном парке «Самаровский Чугас» // Биологические ресурсы и природопользование : сб. науч. тр. Сургут, 2006. Вып. 9. С. 247-253.
8. Москвитин С. С., Москвитина Н. С., Мисуркин О. Н., Павленко Ж. И. Некоторые особенности экологии барсука томского приобья // V съезд всесоюзного териологического общества АН СССР (29 января — 2 февраля 1990 г. , Москва). Москва, 1990.
, Москва). Москва, 1990.
Т. 2. С. 95-96.
9. Новиков Г. А. Теоретические основы и методы изучения питания и трофических связей млекопитающих и птиц // Русский орнитологический журнал. 2001. Экспресс-выпуск 154. С. 673-687.
10.Рудоискатель П. В., Николаенкова А. В., Фадеев К. И. К познанию фауны и некоторых особенностей экологии складчатокрылых ос (Hymenoptera, Vespidae) Свердловской области // Аграрный вестник Урала. 2011. № 2 (81). С. 31-33.
11. Соловьев В. А., Юферев Г. И., Мальцев М. В. Пищевые предпочтения и особенности маркировочной деятельности барсука Волжско-Камского междуречья // Современные проблемы природопользования, охотоведения и звероводства : материалы международной научно-практической конференции, посвященной 85-летию ВНИИОЗ. Киров, 2007. С. 410.
С. 410.
12.Швецов Ю. Г., Смирнов М. Н., Монахов Г. И. Млекопитающие бассейна озера Байкал. Новосибирск : Наука. Сибирское отделение, 1984. С. 88-89.
13.Шилов И. А. Экология. М. : Высш. шк., 1997. 512 с.
14.Hofmann T., Stubbe M. Zur Nahrungsokologie des Dachses Meles meles (L., 1758) in Mitteldeutschland // Beitrage zur Jagd- und
Wildforshung, Bd. 18. 1993. P. 107-119.
15.Goszczynski J., Jedrzejewska B., Jedrzejewski W. Diet composition of badgers (Meles meles) in pristine forest and rural habits of
Poland compared to other European populations // J. Zool. 2000. 250. P. 495-505.
16.Roper T. J., Lups P. Diet of badgers (Meles meles) in central Switserland: an analysis of stomach contents // Z. Saugetierkunde. 1995. 60. P. 9-19.
Saugetierkunde. 1995. 60. P. 9-19.
17. Lanszki J., Kormendi S., Hancz C., Zalewski A. Feeding habits and niche overlap in a Carnivora community of Hungary // Acta Theriologica. 1999. 44 (4). P. 429-442.
18. Middleton A. D. The Food of a Badger (Meles meles) // Journal of Animal Ecology. 1935. Vol. 4. № 2. P. 291.
РАСПРОСТРАНЕНИЕ И СТРУКТУРА
гинекологических заболеваний у коров в племенных организациях свердловской области
М. в. РЯПОСОВА,
кандидат ветеринарных наук, доцент, Уральский НИВИ Россельхозакадемии
Ключевые слова: корова, распространение заболеваний, гипофункция яичников, кисты яичников, органы репродуктивной системы.
Keywords: соw, distribution of diseases, gipofunciya of ovaries, ovarian cysts, organs reproductive system.
В условиях интенсивного ведения молочного скотоводства появляются новые технологии, предъявляющие высокие требования к состоянию здоровья животных. Повышение уровня молочной продуктивности коров сопровождается нарушением обмена веществ, что выражается в ухудшении физиологического состояния и снижении эффективности воспроизводства [1, 2, 3, 4, 5, 6].
Цель исследований.
Изучение распространения и структуры гинекологической заболеваний в племенных хозяйствах при разной технологии содержания животных.
Материалы и методы исследований.
Работа проведена в 2009-2010 гг. в двух сельскохозяйственных организациях Свердловской области с разной технологиях содержания коров: агрофирме «Уральская» (привязная) и ЗАО «Агрофирма «Патруши» (беспривязная).
в двух сельскохозяйственных организациях Свердловской области с разной технологиях содержания коров: агрофирме «Уральская» (привязная) и ЗАО «Агрофирма «Патруши» (беспривязная).
Распространение и структуру гинекологической патологии у коров в сельскохозяйственных организациях при разных технологиях содержания определяли на основании анализа данных учетной и отчетной ветеринарной документации и результатов гинекологической диспансеризации животных. Обследовано 163 бесплодных коровы, не осемененных свыше 45-90 дней после отела или
осемененных, но не оплодотворившихся.
Трансректальное исследование проводили по общепринятой методике. Для подтверждения диагнозов проводили ультразвуковое исследование с использованием портативного УЗИ-сканера «WED-3000» (производство фирмы «Shenzhen Well.D.Electronics Co LTD»). Работа сканера осуществлялась в режиме В, при этом использовался ректальный зонд (датчик) с частотой 7,5 МГ ц (LV2-2/7,5 MHz).
Результаты исследований и их обсуждение.
В исследуемых хозяйствах содержится черно-пестрый скот со среднегодовой молочной продуктивностью 7410 кг и 7759 кг соответственно (табл. 1).
Проведенная ранее диспансеризация коров свидетельствует, что при любой системе содержания животных наблюдаются признаки нарушения обмена веществ. При привязной технологии содержания до 81,28 % животных имеют клинические
признаки нарушения обмена веществ, при беспривязной технологии — 73,26 %. Клинические проявления остеодистрофии и патологии печени регистрируются у 78,11 % и 36,10 % коров при привязном содержании и у 67,07 % и 31,36 % животных при беспривязном содержании [5].
Результаты гинекологической диспансеризации коров в А/Ф «Уральская» показали, что у 52,9 % животных регистрируются заболевания матки воспалительного и невоспалительного характера, среди них ведущее место занимает хронический эндометрит — 41,17 % (у 28 животных). Хроническая субинволюция матки наблюдается у 11,8 % коров (табл. 2).
Хроническая субинволюция матки наблюдается у 11,8 % коров (табл. 2).
Достаточно высокий процент животных регистрируется с кистозными перерождениями яичников — у 17 из обследованных (25,0 %) и еще более высокий процент животных — с гипофункцией яичников (39,7 %). Дисфункции яичников, проявляющиеся фолликулярными и лютеиновыми
Таблица 1
Характеристика племенных хозяйств с разной технологией содержания
Показатели Система содержания коров
А/Ф «Уральская» (привязная) А/Ф «Патруши» (беспривязная)
Численность поголовья коров, гол 1100 1055
Продуктивность, кг 7410 7789
Содержание жира в молоке, % 4,32 4,26
Содержание белка в молоке, % 2,98 3,08
Пищевые привычки перепончатокрылых и Isoptera в тропическом дождевом лесу, согласно соотношению изотопов азота и углерода
Abe T. 1979. Исследования распределения и экологической роли термитов в низинных тропических лесах Западной Малайзии (2) Пища и пищевые привычки термитов в лесном заповеднике Пасо. Яп. J. Ecol. 29: 121-135
Google Scholar
Абэ Т., Бигнелл Д. и Хигаши М. 2000. Термиты: эволюция, социальность, симбиоз, экология . Kluwer Academic Publishers, Дордрехт. 488 pp.
Ambrose S.H. и Норр Л. 1993. Изотопные данные углерода для доставки диетического белка в костный коллаген и из всей диеты в костный апатит-карбонат: эксперименты по выращиванию очищенной диеты. В: Molecular Archeology of Prehistoric Human Bone (Lambert J. and Grupe G., Eds). Шпрингер-Верлаг, Берлин, Германия. pp 1-37
Бейкер Х.и Бейкер И. 1986. Наличие и значение аминокислот в цветочном нектаре. Plant Syst. Evol. 151: 175-186
Google Scholar
Bignell D.E. и Эгглтон П. 2000. Термиты в экосистемах. В: Termites: Evolution, Sociality, Symbiosis, Ecology (Abe T., Bignell D.E. and Higashi M., Eds). Kluwer Academic Publishers, Дордрехт. pp 363-387
Blüthgen N.и Фидлер К. 2002. Взаимодействие между муравьями-ткачами Oecophylla smaragdina , равнокрылыми, деревьями и лианами в пологе австралийских тропических лесов. J. Anim. Ecol. 71: 793-801
Google Scholar
Блютген Н., Гебауэр Г. и Фидлер К. 2003. Распутывание пищевой сети тропических лесов с использованием стабильных изотопов: диетическое разнообразие в сообществе муравьев, богатых видами. Экология 137: 426-435
Google Scholar
Blüthgen N., Gottsberger G. и Fiedler K. 2004. Сахарный и аминокислотный состав нектара и медвяной росы из австралийских тропических лесов. Austral Ecol. 29: 418-429
Google Scholar
Bohart R.M. и Менке А.С. 1976. Sphecid Wasps of the World . Калифорнийский университет Press, Беркли. 600 стр.
Bourguignon T., Šobotník J., Lepoint G., Мартин Дж. М. и Ройзин Ю. 2009. Дифференциация ниши среди неотропических термитов, питающихся почвой, без солдат, выявленная по соотношению стабильных изотопов. Soil Biol. Biochem. 41: 2038-2043
Google Scholar
Брезнак Ю.А. и Брюн А. 1994. Роль микроорганизмов в переваривании лигноцеллюлозы термитами. Annu. Преподобный Энтомол. 39: 453-487
Google Scholar
Брайан М.V. 1978. Производственная экология муравьев и термитов . Издательство Кембриджского университета, Кембридж. 409 pp.
Buschinger A., Klein R.W. и Maschwitz U. 1994. Структура колонии бамбукового жилища Tetraponera sp. (Hymenoptera: Formicidae: Pseudomyrmecinae) из Малайзии. Насекомое. Soc. 41: 29-41
Google Scholar
Дэвидсон Д.В., Кук С.К., Снеллинг Р.Р. и Чуа Т.Х. 2003. Объяснение обилия муравьев в пологах тропических лесов равнин. Наука 300: 969-972
Google Scholar
ДеНиро М.Дж. и Эпштейн С. 1977. Механизм фракционирования изотопов углерода, связанный с синтезом липидов. Наука 197: 261-263
Google Scholar
Де Ниро М.Дж. И Эпштейн С. 1978. Влияние диеты на распределение изотопов углерода в организме животных. Геохим. Космохим. Acta 42: 495-506
Google Scholar
Эгглтон П., Хоматеви Р., Джонс Д.Т., Макдональд Дж.А., Джива Д., Бигнелл Д.Э., Дэвис Р.Г. и Марьяти М. 1999. Сообщества термитов, нарушение лесов и потоки парниковых газов в Сабахе, Восточная Малайзия. Фил. Пер.R. Soc. B 354: 1791-1802
Google Scholar
Фельдхаар Х., Гебауэр Г. и Блютген Н. 2010. Стабильные изотопы: прошлое и будущее в раскрытии секретов питания муравьев (Hymenoptera: Formicidae). Myrmecol. Новости 13: 3-13
Google Scholar
Фиала Б., Машвиц У., То Ю.П. и Хельбиг А.Дж. 1989. Исследования ассоциации муравьев Юго-Восточной Азии — защита деревьев Macaranga с помощью Crematogaster borneensis . Экология 79: 463-470
Fittkau E.J. и Клинге Х. 1973. О биомассе и трофической структуре экосистемы тропических лесов Центральной Амазонки. Biotropica 5: 2-14
Google Scholar
Фрай Б.2006. Экология стабильных изотопов . Спрингер, Нью-Йорк. 308 pp
Gadagkar R. 1991. Belonogaster , Mischocyttarus , Parapolybia и независимое учреждение Ropalidia . В: The Social Biology of Wasps (Ross K.G. and Matthews R.W., Eds). Издательство Корнельского университета, Итака и Лондон. pp 149-190
Gannes L.Z., O’Brien D.M. и Мартинес дель Рио С. 1997. Стабильные изотопы в экологии животных: предположения, предостережения и призыв к проведению дополнительных лабораторных экспериментов. Экология 78: 1271-1276
Google Scholar
Гримальди Д. и Энгель М.С. 2005. Эволюция насекомых . Cambridge University Press, Кембридж, Великобритания 755 стр.
Heil M., Fiala B., Kaiser W. и Linsenmair K.E. 1998. Химический состав пищевых тел Macaranga : адаптация к их роли в привлечении и питании муравьев. Функц.Ecol. 12: 117-122
Google Scholar
Хигаши М. и Эйб Т. 1996. Глобальная диверсификация термитов, обусловленная эволюцией симбиоза и социальности. В: Биоразнообразие: экологическая перспектива (Абэ Т., Левин С.А. и Хигаши М., ред.). Спрингер-Верлаг, Нью-Йорк. стр 83-112
Хобби Э.А. и Вернер Р.А. 2004. Внутримолекулярные, специфичные для соединений и объемные изотопные структуры углерода в растениях C 3 и C 4 : обзор и синтез. New Phytol. 161: 371-385
Google Scholar
Hölldobler B. и Wilson E.O. 1990. Муравьи . Belknap Press издательства Гарвардского университета, Кембридж. 732 pp.
Hyodo F., Kohzu A. and Tayasu I. 2010a. Связывание наземных и подземных пищевых сетей с помощью анализов стабильных изотопов углерода и азота. Ecol. Res. 25: 745-756
Хиодо Ф., Мацумото Т., Такемацу Ю., Камои Т., Фукуда Д., Накагава М. и Итиока Т. 2010b. Структура пищевой сети в тропическом лесу Малайзии на основе соотношений стабильных изотопов углерода и азота. J. Trop. Ecol. 26: 205-214
Хиодо Ф., Таясу И., Иноуэ Т., Адзума Ж.-И. и Кудо Т. 2003. Различная роль симбиотических грибов в деградации лигнина и обеспечении питанием термитов, выращивающих грибы (Macrotermitinae: Isoptera). Функц. Ecol. 17: 186-193
Google Scholar
Хиодо Ф., Таясу И., Конате С., Тондо Дж. Э., Лавель П. и Вада Э. 2008. Постепенное обогащение 15 N с гумификацией рациона в подземной пищевой сети: взаимосвязь между 15 N и возраст диеты, определенный с использованием 14 C. Funct. Ecol. 22: 516-522
Google Scholar
Инуи Ю., Танака Х.О., Хиодо Ф. и Итиока Т. 2009. Обилие тропического таракана в пределах гнезда Pseudoanaplectinia yumotoi , ассоциированного с Crematogaster муравьев, населяющих эпифитный папоротник domatia в борнейском лесу диптерокарпия. J. Nat. Hist. 43: 1139-1145
Google Scholar
Итино Т., Дэвис С.Дж., Тада Х., Хиеда О., Иногучи М., Итиока Т., Ямане С. и Иноуэ Т.2001. Созидание муравьев и растений. Ecol. Res. 16: 787-793
Google Scholar
Джива Д., Бигнелл Д.Э., Эгглтон П. и Марьяти М. 1999. Дыхательный газообмен термитов из скопления Сабах (Борнео). Physiol. Энтомол. 24: 11-17
Google Scholar
Джонс Д.Т. и Гаторн-Харди Ф.Дж. 1995. Собирательная активность процессионного термитника Hospitalitermes hospitalis (Termitidae, Nasutitermitinae) в тропических лесах Брунея на северо-западе Борнео. Насекомое. Soc. 42: 359-369
Google Scholar
Джонс Д.Т. и Престио А.Х. 2002. Исследование термитов (Insecta: Isoptera) района Табалонг, южный Калимантан, Индонезия. Raffles Bull. Zool. 50: 117-128
Google Scholar
Джонс Д.Т., Рахман Х., Бигнелл Д.Э. и Prasetyo A.H. 2010. Леса на ультраосновных почвах Борнео имеют очень бедные ассоциации термитов. J. Trop. Ecol. 26: 103-114
Google Scholar
Кляйн А.М., Штеффан-Девентер И. и Чарнтке Т. 2004. Продолжительность походов за пищей и плотность пчел-мегахилид, ос-эуменид и ос-помпилид в системах тропического агролесоводства. J. Anim. Ecol. 73: 517-525
Google Scholar
Линдеман Р.Л. 1942. Трофико-динамический аспект экологии. Экология 23: 399-417
Google Scholar
Мартин С.J. 1995. Шершни (Hymenoptera: Vespinae) Малайзии. Малайский. Nat. J. 49: 71-82
Google Scholar
Машвиц У. и Мюленберг М. 1975. Стратегия хищничества у некоторых восточных видов Leptogenys (Formicidae-Ponerinae). Экология 20: 65-83
Google Scholar
Мацуура К.1991. Vespa и Provespa . В: The Social Biology of Wasps (Ross K.G. and Matthews R.W., Eds). Издательство Корнельского университета, Итака. pp 232-262
McCutchan J.H., Lewis W.M., Kendall C. and McGrath C.C. 2003. Изменение трофического сдвига для соотношений стабильных изотопов углерода, азота и серы. Ойкос 102: 378-390
Google Scholar
Миченер К.D. 2000. Пчелы мира . Издательство Университета Джона Хопкинса, Балтимор, Мэриленд, 913, стр.
Минагава М. и Вада Э. 1984. Поэтапное обогащение 15 N вдоль пищевых цепей: дополнительные доказательства и связь между δ 15 N и возрастом животных . Геохим. Космохим. Acta . 48: 1135-1140
Google Scholar
Момосе К., Юмото Т., Нагамицу Т., Като М., Нагамасу Х., Сакаи С., Харрисон Р.Д., Итиока Т., Хамид А.А. и Иноуэ Т. 1998. Биология опыления в низинном диптерокарповом лесу в Сараваке, Малайзия. I. Характеристика сообщества растений-опылителей в низинном диптерокарповом лесу. г. J. Bot. 85: 1477-1501
Google Scholar
Надельхоффер К. и Фрай Б. 1994. Исследования изотопов азота в лесных экосистемах. В: Стабильные изотопы в экологии и науке об окружающей среде (Лайта К.and Michener R.H., Eds). Научные публикации Блэквелла, Оксфорд. pp 22-44
O’Neil K.M. 2001. Осы-одиночки: поведение и естественная история . Издательство Корнельского университета, Итака. 406 pp.
Pfeiffer M. и Linsenmair K.E. 2000. Вклад в историю жизни малазийского гигантского муравья Camponotus gigas (Hymenoptera, Formicidae). Насекомое. Soc. 47: 123-132
Google Scholar
Поллирер М.М., Лангель Р., Шеу С. и Мараун М. 2009. Компартментализация пищевой цепи почвенных животных, на что указывает двойной анализ соотношений стабильных изотопов ( 15 N / 14 N и 13 C / 12 С). Soil Biol. Biochem. 41: 1221-1226
Google Scholar
Post D.M. 2002. Использование стабильных изотопов для оценки трофического положения: модели, методы и допущения. Экология 83: 703-718
Google Scholar
Рихтер М.Р. 2000. Пищевое поведение социальных ос (Hymenoptera: Vespidae). Annu. Преподобный Энтомол. 45: 121-150
Google Scholar
Рубик Д.В. 1989. Экология и естественная история тропических пчел .Издательство Кембриджского университета, Нью-Йорк. 514 pp.
Танака Х.О., Инуи Ю. и Итиока Т. 2009. Анти-травоядные эффекты вида муравьев, Crematogaster difformis , населяющего мирмекофитные эпифиты в пологе тропических равнинных тропических лесов на Борнео. Ecol. Res. 24: 1393-1397
Google Scholar
Таясу И. 1998. Использование соотношений изотопов углерода и азота в исследованиях термитов. Ecol. Res. 13: 377-387
Google Scholar
Таясу И., Эйб Т., Эгглтон П. и Бигнелл Д.Э. 1997. Соотношение изотопов азота и углерода у термитов: индикатор трофического образа жизни на градиенте от питания древесиной к питанию почвой. Ecol. Энтомол. 22: 343-351
Google Scholar
Таясу И., Хиодо Ф., Такемацу Ю., Сугимото А., Иноуэ Т., Киртибутр Н. и Абэ Т. 2000. Стабильные соотношения изотопов и сохранение мочевой кислоты у термитов, принадлежащих к трем типам питания в Таиланде. Isotopes Environ. Медицинский Стад . 36: 259-272
Tillberg C.V. и Breed M.D. 2004. Помещение всеядного животного в сложную пищевую сеть: вклад диеты во взрослую биомассу муравья. Biotropica 36: 266-272
Google Scholar
Тилберг К.В., Маккарти Д.П., Долезал А.Г., Суарес А.В. 2006. Измерение трофической экологии муравьев с помощью стабильных изотопов. Насекомое. Soc. 53: 65-69
Google Scholar
Тилман Д. 1982. Конкуренция ресурсов и структура сообщества . Princeton University Press, Princeton, N.J. 296 pp
Van Mele P. and Cuc N.T.T. 2000. Эволюция и статус Oecophylla smaragdina (Fabricius) как средства борьбы с вредителями цитрусовых в дельте Меконга, Вьетнам. Внутр. J. Pest Manage. 46: 295-301
Google Scholar
Вандерклифт М.А. и Понсард С. 2003. Источники различий в рационе потребителей δ 15 Обогащение N: метаанализ. Экология 136: 169-182
Google Scholar
Wäckers F.L. 2004. Оценка пригодности цветущих трав в качестве источников питания паразитоидов: привлекательность цветов и доступность нектара. Biol. Продолж. 29: 307-314
Google Scholar
Ватанабе Х. и Токуда Г. 2010. Целлюлозолитические системы у насекомых. Annu. Преподобный Энтомол. 55: 609-632
Google Scholar
Уилсон Э.О. 1958. Зарождение кочевого и группового хищничества у понеринских муравьев. Evolution 12: 24-31
Google Scholar
Юмото Т. и Накашизука Т. 2005. Программа биологии растительного покрова в Сараваке: объем, методы и достоинства. В: Экология опыления и тропический лес (Рубик Д.В., Сакаи С. и Хамид А.А., ред.). Спрингер, Нью-Йорк. pp 13-21
Hymenoptera (муравьи, пчелы, осы) | ЛОР 425 — Общая энтомология
Основные семьиМухи-кусачки: В большинстве случаев пищу с кровью принимают только взрослые самки.
- Culicidae (комары) — могут переносить малярию, энцефалит, желтую лихорадку, филяриоз и другие заболевания.
- Tabanidae (слепень / олень) — может распространять туляремию, лоаз, трипаносомоз и другие болезни.
- Simulidae (мошки) — могут распространять онхоцериоз человека и лейкоцитозные инфекции домашней птицы.
- Psychodidae (мотылек) — может распространять лейшманиоз, песчаную лихорадку и другие болезни.
- Ceratopogonidae (панки, невидимые глаза) — маленькие, но злобные кусаки, которые были связаны с распространением нескольких круглых червей, простейших и вирусных патогенов среди людей и других животных.
- Muscidae (Комнатные мухи) — одни из самых космополитичных насекомых. У некоторых видов есть кусачий ротовой аппарат, у других — просто падальщики. Такие заболевания, как дизентерия, холера и фрамбезия, могут передаваться через стопы и ротовой аппарат.
Травоядные животные: личинок питаются тканями растений.
- Cecidomyiidae (галлицы) — некоторые вызывают образование галлов растений; другие — падальщики, хищники или паразиты. Это семейство включает гессенскую муху Mayetolia destructor .
- Tephritidae (плодовые мухи) — многие виды являются сельскохозяйственными вредителями; например, яблочная личинка, Rhagoletis pomonella .
- Agromyzidae — большинство личинок — минорцы, некоторые — мотыльки для стеблей и семян.Некоторые виды являются сельскохозяйственными вредителями.
- Anthomyiidae — многие виды являются корневыми или семенными личинками.
Падальщики: личинок питаются навозом, падалью, мусором или другими органическими веществами.
- Drosophilidae (жмыхи) — питаются гниющими плодами.
- Tipulidae (журавлиные мухи) — личинки живут в почве или иле.
- Calliphoridae (мухи) — личинки питаются мусором и падалью; включает червя Cochliomyia hominivorax .
- Chironomidae (мошки) — водные личинки обычно живут в иле и питаются органическими веществами.
- Sarcophagidae (мясные мухи) — личинки обычно питаются падалью. Некоторые виды могут вызывать миаз у человека.
Хищники: взрослых особей и / или личинок нападают на других насекомых как добычу:
- Asilidae (мухи-разбойники) — общие хищники других насекомых.
- Bombyliidae (пчелиные мухи) — хищные личинки; имитирует взрослую пчелу.
- Empididae (танцующие мухи) — взрослые особи хищны.
- Syrphidae (цветочные мухи) — некоторые личинки являются хищниками тлей; большинство взрослых людей имитируют пчел или ос.
Паразиты: личинок являются паразитами или паразитоидами других животных.
- Tachinidae — паразитоиды других насекомых. Некоторые виды являются важными агентами биоконтроля.
- Sciomyzidae (болотные мухи) — личинки паразитируют на слизняках и улитках.
- Oestridae (мухи / камыши) — личинки являются эндопаразитами млекопитающих, в том числе человека.
- Hippoboscidae (вши) — взрослые особи являются кровососущими эктопаразитами птиц и млекопитающих.
- Holometabola
- Полное развитие (яйцо, личинка, куколка, имаго)
- Двукрылых традиционно делят на три подотряда:
- Nematocera (мухи с многочлениковыми усиками)
- Brachycera (летит со стилизованными усиками)
- Cyclorrhapha (мухи с приподнятыми усиками)
- В некоторых более новых классификациях Brachycera включает Cyclorrhapha.
В изобилии по всему миру. Личинки встречаются во всех пресноводных, полуводных и влажных земных средах.
| Северная Америка | Во всем мире | |
| Количество семей | 108 | 130 |
| Количество видов | 16 914 | ~ 98,500 |
В отряд двукрылых входят все настоящие мухи.Эти насекомые отличаются тем, что их задние крылья уменьшены до небольших булавовидных структур, называемых жужжальцами — только перепончатые передние крылья служат аэродинамическими поверхностями. Жужжальцы вибрируют во время полета и работают как гироскоп, помогая насекомому сохранять равновесие.
Все личинки двукрылых безногие. Они живут в водной (пресная вода), полуводной или влажной наземной среде. Они обычно находятся в почве, тканях растений или животных, а также в падалях или навозе — почти всегда там, где опасность высыхания незначительна.Некоторые виды являются травоядными, но большинство питаются мертвым органическим веществом или паразитируют на других животных, особенно позвоночных, моллюсках и других членистоногих. В более примитивных семействах (подотряд Nematocera) личинки мух имеют хорошо развитые головные капсулы с нижнечелюстным ротовым аппаратом. Эти структуры уменьшены или отсутствуют в более продвинутых подотрядах (Brachycera и Cyclorrhapha), где личинки, известные как личинки, имеют червеобразные тела и только пару крючков во рту для кормления.
Взрослые мухи обитают в самых разных средах обитания и демонстрируют огромные различия по внешнему виду и образу жизни.Хотя у большинства видов есть ротовой аппарат, состоящий из двух частей, и они собирают пищу в жидкой форме, их ротовые органы настолько разнообразны, что некоторые энтомологи подозревают, что пищевые адаптации могли возникнуть в результате более чем одного эволюционного происхождения. Во многих семьях хоботок (рострум) приспособлен для обтирания губкой и / или притирания. Эти мухи выживают за счет пади, нектара или экссудатов различных растений и животных (мертвых или живых). В других семьях хоботок приспособлен для разрезания или прокалывания тканей хозяина.Некоторые из этих мух являются хищниками других членистоногих (например, мухи-разбойники), но большинство из них являются внешними паразитами (например, комарами и оленьими мухами), которые питаются кровью своих позвоночных-хозяев, включая людей и большинство диких и домашних животных.
Экономическое значениеДвукрылые, вероятно, оказывают на человека большее экономическое воздействие, чем любая другая группа насекомых. Одни мухи являются вредителями сельскохозяйственных растений, другие передают болезни человеку и домашним животным.С другой стороны, полезны многие мухи, особенно те, которые опыляют цветущие растения, способствуют разложению органических веществ или служат в качестве агентов биологической борьбы с насекомыми-вредителями.
Интересные факты- Несмотря на то, что у них всего два крыла, мухи являются одними из лучших воздухоплавателей в мире насекомых — они могут парить, летать назад, поворачиваться на месте и даже летать вверх ногами, чтобы приземлиться на потолок.
- У мух самая высокая частота взмахов крыльями среди всех животных.У некоторых крошечных мошек он может достигать 1000 ударов в секунду. Самцов комаров привлекает частота взмахов крыльев девственной самки.
- Личинки некоторых прибрежных мух (семейство Ephydridae) живут в необычных местах обитания, которые могут убить других насекомых. Например, Ephydra brucei обитает в горячих источниках и гейзерах, где температура воды превышает 112 градусов по Фаренгейту; Helaeomyia petrolei развивается в залежах сырой нефти; и морская муха, Ephydra cinera , может выдерживать очень высокие концентрации соли.
- Ариста в антенне высших мух — это индикатор воздушной скорости. Это позволяет насекомому ощущать, насколько быстро оно движется.
- По мере взросления куколки мошек надуваются воздухом. При появлении кожа куколки раскрывается, и взрослая муха всплывает на поверхность воды внутри пузыря воздуха. Он никогда не промокает даже ногами!
- Маленькая летучая муха, Megaselia scataris, (Phoridae), действительно всеядное животное. Он был выращен из гниющей растительности, крема для обуви, эмульсий красок, человеческих трупов, маринованных в формалине, и даже тканей легких живых людей.
- Марка комаров с желтой лихорадкой из Габона:
Марка:
Комар желтой лихорадки Марка из Габона:
Комар желтой лихорадки (Aedes aegypti) представляет угрозу для здоровья человека на большей части тропиков. Помимо желтой лихорадки, он также является важным переносчиком вирусов денге и чикунгунья. Эта марка была выпущена Габонской африканской республикой в 1983 году.
Прямые свидетельства питания пыльцой эвдикота у жалящей осы мелового периода (Angiospermae; Hymenoptera, Aculeata), сохранившиеся в бирманском янтаре
Kress, WJ, & Beach, JH в La Selva: экология и естественная история неотропического дождевого леса (ред. Л.А. Макдейд, К.С. Бава, Х.А. Хеспенхейд и Г.С. Хартсхорн), стр. 161−182, 486 (Univ. Chicago Press , 1994).
Momose, K. et al. Биология опыления в низинном диптерокарповом лесу в Сараваке, Малайзия. I. Характеристики сообщества растений-опылителей в низинном диптерокарповом лесу. г. J. Bot. 85 , 1477–1501 (1998).
CAS PubMed Статья Google Scholar
Крепет, В. Л. и Никлас, К. Дж. Вторая «отвратительная загадка» Дарвина: почему существует так много видов покрытосеменных? г. J. Bot. 96 , 366–381 (2009).
Артикул Google Scholar
Гримальди Д. А. и Энгель М. С. Эволюция насекомых (Cambridge Univ.Press, Нью-Йорк / Кембридж, 2005).
Google Scholar
Като М., Иноуэ Т. и Нагамицу Т. Биология опыления Gnetum (Gnetaceae) в низинном смешанном диптерокарповом лесу в Сараваке. г. J. Bot. 82 , 862–868 (1995).
Артикул Google Scholar
Thien, L. B. et al. Биология опыления базальных покрытосеменных растений (сорт ANITA). г. J. Bot. 96 , 166–182 (2009).
PubMed Статья Google Scholar
Luo, S. X. et al. Крупнейшее семейство рано расходящихся покрытосеменных в основном опыляется яйцекладущими насекомыми, как и большинство сохранившихся линий ранних покрытосеменных. Proc. R. Soc. B Biol. Sci. 285 , 20172365 (2018).
Артикул Google Scholar
Labandeira, C. C. Опыление семенных растений среднего мезозоя и ранняя история длиннохоботковых насекомых. Ann. Мисс Бот. Гард. 97 , 469–514 (2010).
Артикул Google Scholar
Рен, Д. Связанные с цветами мухи Brachycera как ископаемое свидетельство происхождения юрских покрытосеменных. Science 280 , 85–88 (1998).
CAS PubMed Статья Google Scholar
Peñalver, E. et al. Длиннохоботковые мухи как опылители голосеменных меловых растений. Curr. Биол. 25 , 1917–1923 (2015).
PubMed Статья CAS Google Scholar
Ren, D. et al. Вероятный способ опыления перед покрытосеменными: евразийские, длиннохоботковые скорпионы. Наука 326 , 840–847 (2009).
CAS PubMed PubMed Central Статья Google Scholar
Гримальди Д. и Джонстон М. А. Длинноязычный меловой скорпион Parapolycentropus Гримальди и Расницын (Mecoptera: Pseudopolycentropodidae): новые данные и интерпретации. г. Mus. Новит. 3793 , 1–25 (2014).
Артикул Google Scholar
Лин, X., Лабандейра, К. К., Ши, К., Хоттон, К. Л. и Рен, Д. Жизненные привычки и эволюционная биология новых двукрылых длиннохоботковых скорпионов из янтаря Мьянмы среднего мела. Nat. Commun. 10 , 1235 (2019).
PubMed PubMed Central Статья CAS Google Scholar
Ларсон, Б. М. Х., Кеван, П. Г. и Иноуэ, Д. В. Мухи и цветы: Таксономическое разнообразие антофилов и опылителей. банка. Энтом. 133 , 439–465 (2001).
Артикул Google Scholar
Красилов В.А., Расницын, А. П. Уникальная находка: пыльца кишечника раннемеловых пилильщиков. Палеонтол. J. 4 , 80–95 (1982).
Google Scholar
Красилов В., Теклева М., Мейер-Меликян Н. и Расницын А. Новый морфотип пыльцы от сжатия кишечника мелового насекомого и его влияние на палиноморфологическую эволюцию и палеоэкологию. Cret. Res. 24 , 149–156 (2003).
Артикул Google Scholar
Вильгельмсен, Л. Предротовая полость нижних перепончатокрылых (Insecta): сравнительная морфология и филогенетическое значение. Zool. Scr. 25 , 143–170 (1996).
Артикул Google Scholar
Красилов В.А., Бугдаева Е.В. Колыбельное сообщество покрытосеменных и новые таксоны проангиоспермов. Acta Palaeobot. Доп. 2 , 111–127 (1999).
Google Scholar
Красилов В.А. Палеоэкология наземных растений. Основные принципы и методы, , с. 283 (John Wiley & Sons, New York, 1975).
Kvaček, J. & Pacltová, B. Bayeritheca hughesii gen. et sp. nov., новый орган, несущий пыльцу Eucommiidites , из сеномана Богемии. Cret.Res. 22 , 695–704 (2001).
Артикул Google Scholar
Аббинк, О. А., Ван Конейнбург-Ван Циттерт, Дж. Х. А., Вишер, Х. Модель экогруппы спороморф для юрского и нижнего мелового периодов Северо-Западной Европы: концепции и основы. Геол. Mijnb. 83 , 17–31 (2004).
Артикул Google Scholar
Лабандейра, К.C. in Эволюционная биология мух (ред. Д. Йейтс и Б. М. Вигманн), стр. 217–273 (Columbia Univ. Press, Нью-Йорк, 2005).
Peñalver, E. et al. Опыление мезозойских голосеменных растений трипсами. PNAS 109 , 8623–8628 (2012).
PubMed Статья Google Scholar
Перис, Д., Пеньялвер, Э., Делкло, Х., Баррон, Э., Перес-де ла Фуэнте, Р., и Лабандейра, К.C. Ложные пузырчатые жуки и распространение способов опыления голосеменных – насекомых до доминирования покрытосеменных. Curr. Биол . https://doi.org/10.1016/j.cub.2017.02.009 (2017).
Cai, Ch., Et al. Опыление саговников жуками в мезозое. Curr. Биол. https://doi.org/10.1016/j.cub.2018.06.036 (2018).
Mao, Y., et al. Различные морские животные с янтарем в бирманском янтаре с обсуждениями его возраста. Палеоэнтомол .https://doi.org/10.11646/palaeoentomology.1.1.11 (2018).
Caldas, M. B., Martins-Neto, R.G., и Lima-Filho, F. P. Afropollis sp. (polén) нет трато кишечника де vespa (перепончатокрылые: Apocrita: Xyelidae) нет Cretáceo da Bacia do Araripe. Атас II Симпос. Nacion. Estud. Tectonico Soc. Бразилия. Геология 195–196 (1989).
Дойл, Дж. А., Джардин, С. и Дёренкамп, А. Afropollis , новый род пыльцы ранних покрытосеменных, с примечаниями о палиностратиграфии мелового периода и палеоокружении Северной Гондваны. Бык. Cent. Recher. Explor. Prod. Эльф. Аквитания 6 , 39–117 (1982).
Google Scholar
Дойл, Дж. А., Хоттон, К. Л. и Уорд, Дж. В. Раннемеловые тетрады, зонасулькатная пыльца и Winteraceae II. Кладистический анализ и последствия. г. J. Bot. 77 , 1558–1568 (1990).
Артикул Google Scholar
Фриис, Э. М., Крейн, П. Р. и Педерсен, К. Р. Ранние цветы и эволюция покрытосеменных растений . стр 585 (Cambridge Univ. Press, Кембридж, 2011).
Пойнар, G. Jr Cascoplecia insolitis (Diptera: Cascopleciidae), новое семейство, род и вид мухи-единорога (Bibionomorpha), посещающей цветы, в бирманском янтаре раннего мелового периода. Cret. Res. 31 , 71–76 (2010).
Артикул Google Scholar
Гримальди, Д. А. и Росс, А. Дж. В Terrestrial Conservation Lagerstätten, Windows into the Evolution of Life on Land (ред. Н. К. Фрейзер и Х.-Д. Сьюз), стр. 287-342 (Данидин, Эдинбург, 2017).
Чжан К., Расницын А. П., Ван Б. и Чжан Х. Перепончатокрылые (осы, пчелы и муравьи) в бирманском янтаре среднего мела: обзор фауны. Proc. Геол. Доц. 129 , 736–747 (2018).
Артикул Google Scholar
Brothers, D. J. Филогения и эволюция ос, муравьев и пчел (Hymenoptera, Chrysidoidea, Vespoidea и Apoidea). Zool. Scr. 28 , 233–250 (1999).
Артикул Google Scholar
Братья Д. Дж. И Карпентер Дж. М. Филогения Aculeata: Chrysidoidea и Vespoidea (Hymenoptera). J. Hym. Res. 2 , 227–304 (1993).
Google Scholar
Расницын, А.П. Hymenoptera-Apocrita мезозоя . Труды Палеонтолог. Instit. Акад. АН СССР 147-1-1321975).
Паклтова Б. Палинологическое изучение покрытосеменных из формации Перук (? Альб – нижний сеноман) Богемии. Sborník Geol. vĕd, Paleontol. Přada 13 , 105–139 (1971).
Уорд, Дж. В. Раннемеловая пыльца покрытосеменных из формаций Шайенн и Кайова (Альб) в Канзасе. США Палеонт. Abteil. В 202 , 1–81 (1986).
Google Scholar
Barrón, E. et al. Палинология аптских и верхнеальбских (нижний мел) янтарных обнажений южной окраины Баскско-Кантабрийской впадины (север Испании). Cret. Res. 52 , 292–312 (2015).
Артикул Google Scholar
Пейро, Д., Barrón, E., Polette, F., Batten, D. J. & Néraudeau, D. Раннесеноманские палинофлоры и предполагаемые смолистые леса и типы растительности в Шаранте (юго-запад Франции). Cret. Res. 94 , 168–189 (2019).
Артикул Google Scholar
Крукшенк, Р. Д. и Ко, К. Геология янтарной местности в долине Хукаунг на севере Мьянмы. J. Asian Earth Sci. 21 , 441–455 (2003).
Артикул Google Scholar
Койро М., Дойл Дж. А. и Хилтон Дж. Насколько глубок конфликт между молекулярными и ископаемыми данными о возрасте покрытосеменных растений? Новый Фитол . https://doi.org/10.1111/nph.15708 (2019)
Крепет, У. Л., Никсон, К. К., Гримальди, Д. и Риччио, М. Мозаичный лауральский цветок из раннего мелового периода Мьянмы. г. J. Bot. 103 , 290–297 (2016).
PubMed Статья Google Scholar
Лю, З.-Дж., Хуанг, Д., Цай, К., и Ван, X. Основной бум эвдикота зарегистрирован в янтаре Мьянмы. Sci. Репу . 8 https://doi.org/10.1038/s41598-018-35100-4 (2018).
Пойнар, Г. О. и Чемберс, К. Л. Palaeoanthella huangii gen. и sp. nov., ранний меловой цветок (Angiospermae) в бирманском янтаре. Сида 21 , 2087–2092 (2005).
Google Scholar
Пойнар, Г. О. и Чемберс, К. Л. Endobeuthos paleosum gen. et sp. nov., ископаемые цветы неопределенного родства из мьянманского янтаря среднего мелового периода. J. Bot. Res. Instit. Tex. 12 , 133–139 (2018).
Google Scholar
Пойнар, Г. О. Programinis burmitis gen. et sp. nov. и P.ламинатус sp. nov., Раннемеловые травянистые однодольные в бирманском янтаре. Austr. Syst. Бот. 17 , 497–504 (2004).
Артикул Google Scholar
Пойнар, Г. О., Чемберс, К. Л. и Бакли, Р. Eoëpigynia burmensis gen. и sp. nov., раннемеловой цветок эвдикота (Angiospermae) в бирманском янтаре. J. Bot. Res. Instit. Tex. 1 , 91–96 (2007).
Google Scholar
Аткинсон Б. А., Стоки Р. А. и Ротвелл Г. А. Отслеживание первоначального разнообразия астерид: анатомически сохраненные плоды корнялина из раннего коньяка (поздний меловой период) западной части Северной Америки. Внутр. J. Plant Sci. 179 , 21–35 (2018).
Артикул Google Scholar
Кеван, П. Г. и Бейкер, Г. Г. Насекомые как посетители цветов и опылители. Ann. Преподобный Энтом. 28 , 407–453 (1983).
Артикул Google Scholar
Джад, Н. А. Ископаемые свидетельства травянистой диверсификации ранних покрытосеменных эвдикотов в раннем меловом периоде. Proc. R. Soc. B Biol. Sci. 282 , 20151045 (2015).
Артикул Google Scholar
Магальон, С., Гомес-Асеведо, С., Санчес-Рейес, Л. Л. и Эрнандес-Эрнандес, Т. Метакалиброванное дерево времени документирует ранний рост филогенетического разнообразия цветковых растений. New Phytol. 207 , 437–453 (2015).
PubMed Статья Google Scholar
Хьюз, Н. Ф. и Макдугалл, А. Б. Барремско-аптские записи пыльцы покрытосермид из южной Англии. Rev. Palaeobot. Палинол. 65 , 145–151 (1990).
Артикул Google Scholar
Чемберс, К. Л., Пойнар, Г. О.И Бакли, Р. Tropidogyne , новый род раннемеловых эвдикотов (покрытосеменных) из бирманского янтаря. Новон 20 , 23–29 (2010).
Артикул Google Scholar
Crepet, W. L. Летопись окаменелостей покрытосеменных: реквием или возрождение? Ann. Мисс Бот. Гард. 95 , 3–34 (2008).
Артикул Google Scholar
Данфорт Б. Н. и Пойнар Г. О. Морфология, классификация и древность Melittosphex burmensis (Apoidea: Melittosphecidae) и последствия для ранней эволюции пчел. J. Paleo. 85 , 882–891 (2011).
Артикул Google Scholar
Гримальди, Д. А. Базальные циклорафа в янтаре мелового и третичного периода (Insecta, Diptera) и их взаимосвязь. Бык. Являюсь.Mus. Nat. Hist. 423 , 1–97 (2018).
Артикул Google Scholar
Менцель Р. и Мюллер У. Обучение и память у медоносных пчел: от поведения к нервным субстратам. Ann. Rev. Neurosci. 19 , 379–404 (1996).
CAS PubMed Статья Google Scholar
Цейл, Дж., Кельбер, А. и Восс, Р. Структура и функция обучающих полетов у наземных гнездящихся пчел и ос. J. Exp. Биол. 199 , 245–252 (1966).
Google Scholar
Джонс, П. Л. и Агравал, А. А. Обучение насекомых-опылителей и травоядных животных. Annu. Преподобный Энтомол. 62 , 53–71 (2017).
CAS PubMed Статья Google Scholar
Фаррис, С. М. и Шульмейстер, С. Паразитоидизм, а не социальность, связан с эволюцией сложных грибовидных тел в мозге перепончатокрылых насекомых. Proc. R. Soc. B Biol. Sci. 278 , 940–951 (2010).
Артикул Google Scholar
Карпентер, Дж. М. и Расницын, А. П. Mesozoic Vespidae. Psyche 97 , 1–20 (1990).
Артикул Google Scholar
Перрард А., Гримальди Д. А. и Карпентер Дж. М. Ранние ветви Vespidae (Hymenoptera) в меловом янтаре. Syst. Ent. 42 , 379–386 (2017).
Артикул Google Scholar
Grimaldi, D. A. Разнообразные орторрафанские мухи (Diptera: Brachycera) в янтаре из мелового периода Мьянмы. Brachycera в янтаре мелового периода, часть VII. Бык. Являюсь. Mus. Nat. Hist. 408 , 131 (2016).
Артикул Google Scholar
Фейлд Т.С., Аренс, Н. К., Дойл, Дж. А., Доусон, Т. Э. и Донохью, М. Дж. Темные и обеспокоенные: новый образ экологии ранних покрытосеменных. Палеобиол 30 , 82–107 (2004).
Артикул Google Scholar
Гримальди Д. Совместное излучение насекомых-опылителей и покрытосеменных в меловом периоде. Ann. Мисс Бот. Гард. 86 , 373–406 (1999).
Артикул Google Scholar
Shi, G. et al. Ограничение по возрасту бирманского янтаря на основе U-Pb датирования цирконов. Cret. Res. 37 , 155–163 (2012).
Артикул Google Scholar
Пунт В., Хоэн П. П., Блэкмор С., Нильссон С. и Ле Томас А. Глоссарий терминологии по пыльце и спорам. Rev. Palaeobot. Палинол. 143 , 1–81 (2007).
Артикул Google Scholar
Hesse, M. et al. Терминология пыльцы. Иллюстрированный справочник . С. 266 (Springer, Вена, 2009).
Schindelin, J. et al. Фиджи: платформа с открытым исходным кодом для анализа биологических изображений. Nat. Методы 9 , 676–682 (2012).
CAS Статья Google Scholar
Шинделин, Дж. И др., Экосистема ImageJ: открытая платформа для анализа биомедицинских изображений. Мол. Репродукция. Развивать. PMID 26153368 (2015).
Preibisch, S., Saalfeld, S. & Tomancak, P. Оптимальное сшивание мозаичных трехмерных изображений, полученных с помощью микроскопа. Биоинформатика 25 , 1463–1465 (2009).
CAS PubMed PubMed Central Статья Google Scholar
Федоров А. и др. 3-D Slicer как платформа обработки изображений для сети количественной визуализации. Magn. Резон. Представь. 30 , 1323–1341 (2012).
Артикул Google Scholar
Филогения APG-покрытосеменных, группа IV. Обновление классификации филогенетической группы покрытосеменных для порядков и семейств цветковых растений: APG IV. Бот. J. Linn. Soc. 181 , 1–20 (2016).
Артикул Google Scholar
Личиночное питание гемолимфы и обнаружение гемолимфы у муравья Proceratium itoi (Hymenoptera: Formicidae) | Masuko
Личиночное питание гемолимфы и выделение гемолимфы у муравьев
Proceratium itoi (Hymenoptera: Formicidae)Абстрактные
Королевы видов муравьев Proceratium itoi (Forel, 1918) обычно получают питание посредством явления, известного как личиночное питание гемолимфой (LHF), когда они питаются гемолимфой личинок последнего (4-го) возраста своего потомства. .Каждая личинка имеет четыре пары специализированных структур на дорсальных покровах со 2-го по 5-й брюшные сегменты. Каждая структура состоит из небольшой кутикулярной области с мелко потрескавшейся поверхностью, напоминающей поверхность кукурузного початка. Из-за этих трещин царица может вскрыть кутикулярную поверхность защемлением нижней челюсти и поглотить гемолимфу из образовавшихся отверстий. Эту структуру можно рассматривать как ответвление личиночной гемолимфы P. itoi и является вторым подтвержденным случаем такого органа у муравьев (первый был у Leptanilla ).Королева в развитой колонии зависит исключительно от гемолимфы личинок для своего питания, но в зарождающейся колонии с небольшой популяцией королева подавляет LHF и питается только добычей. Между тем рабочие питаются добычей и редко выполняют LHF, за исключением случаев, когда колония голодает. Раны на покровах личинок закрываются вскоре после LHF за счет коагуляции гемолимфы, и личинки неоднократно подвергаются LHF, демонстрируя множественные шрамы, характерные для LHF, на их спине. Подобные рубцы также обнаруживаются на спинке многих предкуколок, что указывает на то, что они могут выжить при LHF и созреть.
Открытый доступ, под лицензией CC BY 4.0. © 2018 Автор (ы)
DOI: https://doi.org/10.25849/myrmecol.news_029:021
Страницы: 21-34
Том: 29
Год: 2019
Журнал: Myrmecol. Новости
Ключевые слова
Муравей, Proceratium itoi, матка, личинка, питание гемолимфой, выделение гемолимфы, коагуляция.
Файл:
(Контакт)
перепончатокрылые Опылители
Порядок Перепончатокрылые входит в четверку крупнейших отрядов насекомых, насчитывающую более 300 000 идентифицированных разновидность.Однако оценки общее количество существующих видов превысило один миллион. Орден характеризуется наличием двух пары перепончатых крыльев с сетчатыми жилками, состоящими из 20 или менее клетки. Губная и верхняя челюсти объединены в структуру всасывания и притирания. Челюсти хорошо развиты для жевания и захвата добычи. Стремительное и подвижное тело сильно бронированный. Самки откладывают яйца структура (яйцеклад), разработанная для прокалывания.Различают стадии личинки и куколки (= голометаболический). Личинки земные и могут быть как активными, так и вырожденными. Взрослые кормят своих детенышей, откладывая яйца в источник пищи или путем хранения продуктов. Взрослые особи большинства видов хотя бы частично питаются пыльцой и нектар. Есть три основные группы в таком порядке: Кормление растений, Паразитарный и жалящий.
Группа растениеводства (Симфита).
Пилильщики, роговицы и груши слизняк являются примерами. Яйцеклад пилообразный, брюшко и грудная клетка широко соединены. Личинки имеют 13 члеников, хорошо развиты. грудные ноги и обычно несколько пар брюшных ложноножек. Они питаются живой тканью растений. Очень немногие виды паразитируют. Взрослые особи одной семьи, Tenthredinidae (пилильщики) часто встречаются на цветках, особенно II и III групп .Они преобладают на лютиках в больших количествах. каньоны Скалистые горы Северной Америки. Личинки некоторых виды пилильщиков строят галлы на ивах. Несколько видов были замечены в большом количестве опыляющих цветет клубника. Тем не менее гусеницы личинки руды очень губительны.
Паразитическая группа.
Примеры этой группы включают Ichneumonoidea , Chalcidoidea и Cynipoidea (галловые осы).Яйцеклад обычно длинный и разработан для пирсинга. Яйца проходят по всей длине яйцеклада. Вертлы у большинства родов делятся на два кажущихся сегмента. У личинок нет грудных ног и являются внутренними паразитами (некоторые также являются внутренними паразитами) других насекомых и пауки. Однако желчная оса личинки образуют галлы в тканях растений, а некоторые хальциды заражают семена. Взрослые менее 10 процентов населения виды несколько регулярно питаются цветами, особенно в группах II и III .Но некоторые специально модифицированные члены группы Group VI могут быть использовал. Большинство посетителей цветов находятся в семейство Ichneumonidae. Взрослые могут можно найти на оленьих кустах, бузине, зонтиках, лютиках и сервисберри. Они важны в качестве опылителей небольшого количества цветков ихневмонии Группа VI . Они, вероятно, считают пчел полезными насекомые из-за их паразитических привычек на вредных насекомых.
Жалящая группа (Aculeata).
Муравьи, пчелы, шершни и осы члены этой группы насекомых. В яйцеклад обычно развивается как жалящая структура, снабженная сосуд для яда. Яйца не пересечь яйцеклад. Живот и грудная клетка разделены определенным сужением, а вертлуги не разделенный.У личинок нет ног и обладать разнообразными диетами. Большинство питаются парализованных насекомых или на пыльце и меде. Несколько групп являются внешними паразитами насекомых и две группы — паразиты. муравьи и бумажные осы) всеядны. Взрослые особи большинства родов много времени проводят на цветах и питаются пыльца и нектар. Одна большая группа, пчелы и некоторые другие также собирают пыльцу и мед для своих потомство. Самки большинства виды способны ужалить в целях самозащиты.Это, безусловно, самая ценная из трех основных групп.
У Formicidae (муравьи) основание живота двойное перетяжка, в центре которой находится припухлость или узел. Самки могут встречаться в нескольких повязках. включая бескрылых рабочих и обычно также бескрылых солдат, как с сильно уменьшен грудной отдел. В первый сегмент антенны почти такой же длины, как и все остальные сегменты комбинированный.Корпус отполирован и голые или очень редко волосатые. В почти все личинки всеядны, но у некоторых видов может быть специализированная диета. например, грибок, семена или падь. Все виды муравьев колониальны, и большинство из них развивают очень большие размеры. колонии со многими специализированными кастами. Лишь несколько видов посетят цветы. Те, кто путешествуют в основном пешком, чтобы Обычно клеймо не попадает в цветок. Если бы они действительно добрались до цветка, было бы только самоопыление. удавшийся.Иногда они будут стоять на страже цветка, отгоняя более эффективных опылителей и тем самым быть вредным.
Веспоид Осы включают бархат муравьи, шершни, чешуйчатые осы и осы-кукушки. Задняя доля переднегруди лежит рядом с тегулой. или шапочка у основания переднего крыла. У самого большого семейства, Vespidae, передние крылья складываются вдоль. В нескольких семьях есть бескрылые самки, но Среди социальных веспидов крылья есть у всех каст.Рацион личинок разнообразен и разнообразен. все потребляют пищу, которую дают взрослые женщины. У большинства родов есть личинки, которые питаются парализованными жертвами насекомых, которые обеспечивается материнской осой, но некоторые из них являются внешними или внутренними паразиты. Некоторые виды питаются пыльца, мед и социальные веспиды обычно всеядны. Несколько родов Vespidae являются социальными. и построить маленькие или большие бумажные гнезда. Взрослые особи двух семейств: Scoliidae и Vespidae , часто встречаются на цветках, где они питаются нектаром.Чаще всего используются цветы из Group II и некоторых короткие венчики VI группы . Одна семья (Masaridae) поставляет свой выводок клетки с пыльцой и нектаром на манер пчел. Однако диапазон его хостов очень ограничен. Их можно найти на Phacellia на высоте более 6000 футов в западные горные каньоны. Они появились как большие желтые куртки с булавовидными усиками. Некоторые из более крупных Scoliidae ( Campsomeris ) может споткнуться и опылить люцерну в теплых местах.Потому что они парализуют личинок жука-скарабея для своего потомства, они очень полезны. Их большой волосатое тело хорошо приспособлено для опыления и обычно встречается на поля хлопка и донника.
Помпилиды (Осы-пауки) соток сине-черный или зелено-черный по окрасу, с коротко остриженной шелковистой шерстью. Большинство видов стройные, по бокам сжатые длинные колючие и гладкие ноги, которые создают вид обтекаемые тела.Задний мочка переднегруди почти не касается тегулы. У обоих полов есть крылья. Практически все виды дают потомству парализованные пауки. Они все полностью одиночные насекомые. Взрослые большинства виды питаются падевой росой, выделяемой тлями и щитовками, а также обнаженный нектар в цветах. Они частые цветки тамарикса, ивы, молочая, дикой гречихи и зонтичных. Их ценность как опылителей на самом деле невелика. жизненно важно, потому что цветы, которые они посещают, также привлекательны для многих пчел и осы.
Sphecidae (копатель ос) включить ястребы-цикады, грязевые мази, песчаные осы и осы с нитевидной талией. Задние доли переднегруди не доходят до тегул и нет разветвленных волосков на теле. Самки большинства видов имеют длинную челку. или граблями на передних лапках, которые используются для зачерпывания почвы. У большинства видов лицо серебристого цвета. и у многих наблюдается длинное сужение в основании живота.Сфециды почти всегда обеспечивают потомство со специализированной диетой из парализованных насекомых или пауков. Колониальных видов нет, но есть очень общительный. Взрослые большинства виды питаются нектаром или одновременно пыльцой и нектаром цветов. Эти цветы обычно входят в Группу II, некоторые из группы VI, такие как донник, и некоторые социальные цветы, такие как куст кролика и тысячелистник с короткими трубками венчика. Некоторые из них развили длинные языки и могут получать пищу от многих видов социальных цветов и цветков перепончатокрылых.Одна группа ( Philanthus ) Обеспечивает свои гнезда пчелами. Sphecidae более ценны, чем предыдущие группы перепончатокрылых. как опылители, но не так ценны, как пчелы. Некоторые песчаные осы посещают люцерну и случайно спотыкаются о ней. их ноги, но сомнительно, что это часто приводит к перекрестному опылению. Они одни из самых частых посетителей к сельдерею, моркови, хлопку, доннику, подсолнечнику, авокадо и дикорастущим растениям такие как кроличий куст, золотарник, спичечная неделя и кротон.Увеличить их количество было бы сложно, потому что для этого требуется добыча насекомых. Песчаные районы наиболее благоприятны для большое разнообразие спехид.
Апоидея (Пчелы) состоит более 12 000 видов. Включены пчелы, пчелы пота, шмели, пчелы без жала, пчелы-плотники и кочевники пчелы. У всех видов есть первые предплюсневой сегмент задней ноги не менее двух третей длины голени и у большинства видов она не более чем в три раза длиннее ширины.У всех Apoidea есть несколько разветвленных волосков. их тела. Самки всех, кроме Hylaeus и паразитарных групп имеют отчетливые структуры для сбора пыльцы на брюшке или задних лапах. Большинство видов очень мохнатые или пушистые и у многих длинные выдвижные языки. А характерной привычкой в биологии всех пчел является то, что молодняк нектар и пыльца, собранная с цветов. Лишь несколько родов являются колониальными, но к ним относятся некоторые из самых обильный, такой как пчелы, шмели и некоторые пчелы пота.Многие роды имеют стадные виды. Однако большинство из них совершенно одиноки. Многие роды откладывают яйца на запасы пыльцы других пчел, но в большинстве случаев их не так много. Включены роды Nomada , Psit h yrus, Sphecodes и Coelioxys . Взрослые особи всех видов питаются нектаром и пыльца в дополнение к их собирательной деятельности.За исключением пчел-самцов, пчелы-самцы посещают цветы. для кормления, как и их самки. Однако самки, потому что они также собирают для своих детенышей, более трудолюбивы и постоянны в своем внимании к одному виду цветок, чем самцы. женский пчелы также строят гнезда и ячейки для расплода, хранят корм, откладывают яйца и ухаживать за молодняком. Пчелы характерные места гнездования, охватывающие широкий спектр местообитаний, таких как почва, древесина и полости в полых бревнах, раковины улиток, мышиные гнезда и т. д.).
Apoidea определенно важнейшие агенты опыления. Медоносные пчелы являются наиболее важными в сельскохозяйственных областях. Однако на некоторых участках и для некоторых культур это не тот случай. В дикой природе или на малокультурных территориях есть виды местных пчел, которые могут быть основные опылители как сельскохозяйственных культур, так и дикорастущих растений. Некоторые растения, такие как Phlox и Delphinium , монашеские, ирисы, орхидеи и др., должны зависеть от диких опылителей, потому что пчелы не могут используйте их правильно. Пчелы из-за их широкого диапазона хозяев, долгого сезона, большого размера и того факта, что они контролируются людьми в больших количествах для продуктов их ульи, имеют наибольшие возможности для специфического использования в качестве опылители. Некоторые из родных тем не менее, виды больше подходят для опыления некоторых культур и для определенные климатические условия и в некоторых случаях могут сравниться по количеству с пчелы.Также существует значительный доказательства того, что некоторые из них больше подходят для контролируемого опыления в заключение, например, в теплицах, клетках и т. д. Возможность использования любых особых преимуществ родного пчелы или просто использовать их в качестве дополнения к медоносным пчелам — это хорошо для некоторых ситуаций.
Апоидея классифицирована согласно Биологический тип
Кукушка Пчелы (Anthophoridae).
Эти пчелы откладывают яйца в гнезда других пчел. Их личинки затем потребляйте пыльцу и мед, которые хранились для личинок хозяина. В роду Psithyrus (шмели-гости), взрослая самка занимает гнездо шмеля-хозяина и питается мед хранится хозяином. Она тогда откладывает яйца в клетки, построенные и предоставленные ее хозяевами.Эти яйца высиживают хозяева и, при вылуплении личинки растут как члены семьи! У родов Nomada , Stelis , Melecta и Sphecodes , взрослая самка входит в гнездо, пока оно готовится, и кладет яйцо рядом с яйцом хозяин до того, как ячейка запечатана. Личинка первого возраста поедает яйцо хозяина или молодые личинки, а затем питается хранящимися там медом и пыльцой.
Самки кукушки отличаются отсутствием собирающих пыльцу структур, их редкостью или очень короткое опушение, тяжелая броня и хорошо развитое жжение. аппарат. Расцветки многих видов яркие и контрастные. В некоторых случаях они могут быть тесно связаны со своими хозяевами.
Одиночные пчелы (Andrenidae).
Самки обеспечивает отдельные клетки в собственном гнезде и откладывает одно яйцо на поверхности магазины перед запечатыванием ячейки. Личинка развивается самостоятельно, питаясь хранящейся пищей. Эти пчелы совершенно одиночки. Гнезда не собираются, но имеют тенденцию быть разбросанными по доступным местам гнездования. Они стадные, а гнезда сгруппированы в крупные или мелкие, плотные или рыхлые агрегаты.Между женщинами нет сотрудничества.
Социальные Пчелы (Halictidae).
Сотрудничество между родителями и потомством — это основной шаг к настоящему насекомому общество. В типе Halictus женщина удобряют в конце лета; она зимует и начинает строительство ее гнездо весной точно так же, как пчела-одиночка.Но откладывает только женские яйца. Вместо того, чтобы умереть, когда гнездо будет построено, она останется в нем до тех пор, пока выводок появляется. Молодой новый самки немного меньше своей матери и не имеют самцов, с которыми спариваться. Хотя они кладут какой-то самец яйца, большинство новых клеток, которые они строят в старом гнезде, содержат яйца которые были заложены их матерью. В мать не продолжает добывать корм, но служит яйцекладкой и защитник гнезда.Следующий поколение содержит самцов, которые спариваются с самками того же поколение. Затем эти самки несут виды на следующую зиму. В этом обществе нет прогрессивного кормления молодняка и нет нет сотрудничества между сестрами: только между матерями и дочерьми.
В социальных сетях типа Шмель пчела, поведение аналогично Halictus тип.Однако молодняк кормится постепенно перезимовавшими королева и позже сменяющие друг друга выводки рабочих. В этом случае при постепенном кормлении яйцо откладывается в пустую ячейку или изначально не обеспечен достаточным кормом для вылупившихся личинок до зрелости. Первые пчелы выращенные королевой, низкорослые из-за недостатка пищи, но они предполагают обеспечение обязанностей для последующих поколений, которые все производятся из яйца, отложенные королевой.Рабочие сотрудничать в добыче, кормлении, хранении пищи, уборке и охране гнездо. Последующие поколения лучше кормят, и самцы и новые матки производятся в финальном поколение. Кроме того, как и в случае с типом Halictus , новый оплодотворенная матка должна переносить этот вид всю зиму.
Пчелы без жала Apidae Meliponini (Melipona & Trigona).
Это важная тропическая группа, которая отличается от шмелей главным образом тем, что матка многолетняя, а колония воспроизводится группа рабочих, которая кишит только что произведенной королевой. Колонии вырастают до больших размеров, конкурировать с пчелами, и хранение меда может быть почти таким же Отлично. Королеве не нужно кормить или работать в гнезде, кроме откладывания яиц.Этот тип не кажется таким продвинутым, как шмель. тип. Молодняк не кормится постепенно, но каждая камера снабжается полным набором пищи и запечатанный. Многие люди принимают участие совместно в процессе. Этот тип — это специализированная гостиница, в которой мужчина — настоящий трутень, и его нужно кормить в гнездо рабочими.
Медоносная пчела Тип — Apidae.(Домашние пчелы).
Сюда входят несколько видов из Apis , из них Apis mellifera является членом. Это общество объединяет достижения шмелей и пчел без жала, но есть также несколько дополнительных достижений. Колония размножение — роение, но рабочие вместо этого роятся старой маткой. нового.В этой системе колония никогда не должна вымирать, пока существуют яйца, чтобы рабочие могли развиваться в новый маток специализированным кормлением. Безжалостным пчелам, обитающим в тропиках, не приходится перезимовать. Медоносные пчелы не переживают зиму. впадают в спячку, но из-за сгущения тепла и употребления меда. Шмели и пчелы без жала либо уничтожают старые клетки или используйте их только для хранения меда. Медоносные пчелы могут многократно использовать клетки для хранения пищи. и выращивание выводков.
Опыление Пчелы
Для технических культур в умеренном климате В климатических условиях пчелы являются наиболее ценными насекомыми-опылителями. У медоносных пчел есть ряд общих характеристики хорошего опылителя. Они полностью зависят от цветов в еде и часто только те части растений, которые несут пыльцу или нектар.Они одеты в перистые волосы, которые удерживать пыльцу. Они не вредны растениям и не доставляют неудобств публике. Потому что они так старательно доставляют и хранят нектар и пыльцу для своего потомства, они более надежные посетители цветов, чем насекомые, которым нужно только себя прокормить.
Медоносные пчелы также обладают несколькими чертами что особенно квалифицирует их как опылителей. Некоторые из них характерны и для других пчел, но не для других видов. имеет столько же таких атрибутов.В кормовой сезон медоносных пчел начинается с первых цветков весной и заканчивается последними цветками осенью. Колониями пчел можно в значительной степени управлять, и они могут при необходимости можно перемещать в другие посевные площади. Размер их тела и длина хоботка промежуточные. и дать им возможность работать как с большими, так и с маленькими цветами. Однако некоторые цветы могут быть только эффективно опыляется более крупными или мелкими видами. Из-за необходимости хранить большие продукты излишков, чтобы в активном состоянии пережить зиму необходимо они посещают больше цветов, чем другие виды, которые остаются неактивными для части год.Постоянство индивидуальная медоносная пчела на вид растения увеличивает ее опыление эффективность. Эта черта меньше выражен у многих видов диких пчел. Способность разведывать и собирать медоносных пчел для передачи расположение привлекательных цветов позволяет колонии быстро найти пищу источник. Диапазон хоста пчелы очень широки для одного вида. Таким образом, он может служить опылителем для самых разных растения.
Конечно, у пчел есть некоторые ограничения. как опылители. Они не очень много летают в сырую или пасмурную погоду или при температуре ниже 60 градусов Фаренгейт. Полет сокращен на ветер 10 миль в час и почти прекращается ветрами более 15 миль в час. Таким образом, некоторые дикие пчелы, например шмели, может летать в более неблагоприятных условиях. Медоносные пчелы склонны ограничивать добычу пищи одним районом, и это может приводит к большей тенденции к самоопылению деревьев или клонов. посадки.Они могут получить нектар некоторых видов цветов, например люцерны, без опыления. Кроме того, они могут собирать пыльцу с растения, которые обычно опыляются ветром и силой тяжести, такие как сорго, Рогоз, кукуруза, самшитая бузина и бермудская трава.
Выбор цветов от Медоносные пчелы
Даже если пчелы переедают широкий спектр видов цветов, они, как правило, сосредотачиваются на цветках, которые для них более привлекательны.В концентрация сахара и объем нектара являются важными аттрактантами. Эти атрибуты различаются в зависимости от вида цветка. и разнообразие. Концентрация сахара колеблются от 5 до 80 процентов, но те, кто ниже 40 процентов, менее привлекательны для пчелы. Когда растения выращивают для высокая урожайность семян, селекционеры могут увеличить нектар концентрация сахара путем отбора. Влажность воздуха и время суток также влияют на частоту посещение цветов пчелами.
Когда пыльца в большом количестве Цветочные виды медоносных пчел будут больше привлечены. Они также могут отдавать предпочтение различным видам пыльцы. Пчелы быстро учатся, когда цветы откроют свою пыльцу. Пчелы могут организовать свою полевую деятельность, посылая разведчики. Обычно это люди который выполнял другие обязанности в колонии. Скауты будут искать новые источники пыльцы и нектара и не подвержены влиянию общение с другими пчелами.Когда обнаружен новый источник, они берут образец в улей и сообщают его расположение фуражиров в улье.
Медоносные пчелы — обычно особи. которые имели предыдущий опыт работы в качестве разведчиков. Они посещают источники пищи, которые были переданы в улье разведчики и другие собиратели. В некоторых виды цветов нектар нельзя собирать отдельно от пыльцы, но почти во всех случаях пыльцу можно собрать отдельно от нектара.Но большинство сборщиков пыльцы берут небольшие количество нектара. Некоторый возможно совпадение этой привычки, есть определенное различие. Было замечено, что нектар сборщики обычно более старые пчелы, чем сборщики пыльцы.
Пчелы, возвращающиеся с поля, передают несколько видов информация для других пчел в улье. О том, какой цветок, говорит запах груза. Расстояние и направление к источнику передается сигнальным танцем.Эта информация быстро распространяется по мере того, как информированные пчелы сами находят источник и возвращаются, чтобы стать более информаторы.
Дальность полета до источника питания могут достигать примерно 10 миль, но они предпочитают добывать корм в пределах 1,5 миль от источники пищи многочисленны. В пределах одной мили желательность источника пищи наиболее высока. важный. Молодые полевые пчелы обычно ограничиваются добычей пищи рядом с ульем в течение примерно двух дней.Предпочтительный ассортимент сборщиков пыльцы обычно короче, чем у сборщиков нектара. Для промышленного производства меда пчеловоды обычно не ожидайте, что источник цветка находится на расстоянии более двух миль от улья. При опылении ближе цветок тем больше пчел найдут его и тем больше посещений можно будет сделать.
Количество цветов, которые посещение медоносных пчел зависит от обилия пыльцы или нектара в цветок.Это может быть до 1000 цветы, посещенные за груз нектара в цветах клена, или всего за один из тюльпанного тополя. Это может потребовать как около 400 посещений для груза пыльцы желтого донника или всего лишь 25 посещений из одуванчика. Пчелы редко будет получать полный груз из дефицитных источников. Цветы с максимальным количеством нектара оптимальны для производства меда, но не дают эффективное использование опылителей.
Связь в Пчелы
Медоносные пчелы общаться с другими членами улья, выполняя Хоровод и Виляние Танец .Хоровод обычно исполняется в 50 футов улья и имеет переменную продолжительность. В танце виляния пчелы делают полукруг, а затем бегут в направление источника питания. Это все сделано в соответствии с положением солнца. Количество поворотов в прямом беге передает расстояние. Чем ближе источник пищи тем больше покачиваний в секунду. О типе пищи сообщают образцы, принесенные в улей. Такое поведение характерно только для рода Apis .Некоторые примитивные пчелы используют платформу снаружи улей и направьте прямо на источник пищи. Когда пчелиная семья перемещается с севера на юг Полушарие, это запутает их коммуникативные способности.
Сельское хозяйство Химия и опыление
Агрохимикаты Основное беспокойство в отношении опыления насекомых вызывают фунгициды, гербициды, удобрения, гормоны завязывания плодов, соединения для прореживания цветков и инсектициды и митициды.
Гормоны для завязывания фруктов с рисунком после того, как природные соединения, присутствующие в пыльце, создают стимул для окружающей среды плоды, либо с нашим без опыления. Когда эти гормоны применяются раньше может произойти опыление, партеногенетическое развитие бессемянных плодов. Если они вносятся после опыления, набор плодов с семенами может быть более плодовитым за счет меньшего опадания цветков или плоды могут стать больше, чем обычно.Для некоторых фруктов гормоны, применяемые после формирования плодов, уменьшаются. предуборочная фруктовая капля. В разведении работа с самостерильными растениями, некоторые гормоны могут временно преодолеть самостоятельность. Для коммерческого томаты производства очень успешно обрабатываются гормонами с помощью производство бессемянных и сеянных томатов в теплицах.
Растения, получающие полноценное питание как правило, лучше всего способны обеспечить хорошее опыление, и, следовательно, применение коммерческое удобрение выгодно.Некоторые культуры могут иметь тенденцию к вегетативный, а не репродуктивный рост, если азот-фосфор-калий соотношение не приспособлено к их требованиям. Также существуют различные добавки к удобрениям, например, минералы, которые может повысить привлекательность растений для опылителей.
Химикаты для прореживания цветков, например как динитрос и гормоны, особенно используются в яблоневых садах. Может возникнуть переизбыток завязи фруктов. в яблоках меньшего размера.Это лучше делать химическое прореживание только тогда, когда приближается цветение и опыление оптимальный. С динитрос применение должно быть сделано, когда постукивание по ветке приводит к падению лепестков с дерево. Динитрос и гормоны делают не оказывает вредного воздействия на пчел даже при применении во время дневное время.
Гербициды, применяемые как дефолиаторы или гормоны не оказывают значительного вредного воздействия на пчел, если они не применяются непосредственно к ним.Основное неблагоприятное воздействие таких химикатов на пчел заключается в следующем: уничтожают их корм. Много пыльцы и Медоносные растения важны для медоносных и диких пчел. Однако селективные гербициды могут увеличить обилие ценных кормовых растений пчел, если их использование приводит к сокращение конкурентоспособных видов сорняков.
Фунгициды очень мало известные вредные воздействия на медоносных и диких пчел.
Инсектициды и митициды необходим для борьбы с вредными насекомыми и клещами в коммерческих производство многих сельскохозяйственных культур. В химическая борьба с насекомыми может увеличить корм для медоносных пчел и позволяет растения, чтобы воспользоваться полученным опылением. Однако почти все инсектициды токсичны. пчелам, чтобы избежать применения средств борьбы с насекомыми на стадии цветения, который наиболее привлекателен для пчел.Важно не допускать попадания химикатов на под прикрытие. растения (например, в садах) или дрейфующие к обочине дороги и выбрасывающие цветы или соседние поля в цвету. Бедные методы применения могут привести к гибели пчел-фуражиров, и хотя это может не привести к гибели колонии, это может помешать выращиванию меда и разрушить колонии для эффективного опыления. Последующей проблемой может стать перезимовка колонии. Пчелы-медсестры могут погибнуть, получив отравление материалы от полевых пчел.В широкое распространение соединений на основе мышьяка в начале 20-х гг. -е гг. г. Век был губительным для пчел. Загрязнение меда полевыми химикатами не было Однако это серьезная проблема для потребителей.
При применении инсектициды для цветущих растений не требуются, лучше всего удалить все ульи из непосредственной близости. Из конечно, приложения лучше всего делать, когда пчелы не в поле или фруктовый сад.Также инсектициды с По возможности следует выбирать наименьшую токсичность для пчел.
Обычно используемые инсектициды могут быть оценены по их влиянию на пчел. Некоторые материалы не оказывают заметного воздействия на пчел, если их нанести. пока пчел нет в поле. Это ротенон, пиретрум и никотин, а также сера, масла, фумиганты и динитрос. В современном развитом сельском хозяйстве более доступные по цене инсектициды могут тем не менее, имеют приоритет.По ул. 21 Век многие соединения, которые были токсичны для пчел, были удалены из рынок, в том числе хлорированные углеводороды. Тем не менее, использование некоторых запрещенных материалов может быть широко распространен в странах третьего мира.
Использование диких пчел для Опыление
Дикие пчелы имеют большой потенциал для коммерческого опыления, но их изобилие сильно тормозили рост городов, уничтожение естественного хозяина растений и землепользования, включая дренаж, обработку почвы, орошение, очистку и чистое выращивание.Эрозия, наводнения, использование инсектицидов и гербицидов на цветущих растениях также неблагоприятно для диких популяции пчел.
Тем не менее, есть способы какие дикие пчелы могут быть предпочтительнее. Эти включать введение новых видов меда и пыльцы, дренаж и орошение, благоприятное для популяции пчел, ограждения и здания, покрытые черепицей, отверстия для гвоздей, сараи и т. д. интродукция некоторых полых растений с острым стеблем, таких как расторопша, Ailanthus и т. д.также одобряют дикие пчелы. Все эти изменения часто нестабильны и могут не благоприятствовать популяциям диких пчел в течение длительного времени запустить. Их создание в районе может потребоваться несколько лет.
Некоторые методы консервации могут приводит к созданию популяций диких пчел. Уход за живой изгородью содержательной, растения с полым стеблем, которые были затоптаны, дают диких пчел среда обитания.Естественная растительность должна быть поддерживается на некотором расстоянии по обе стороны от водотоков. Хорошая пыльца и нектар растений должны быть предпочитают на обочинах дорог и на неиспользуемых полях. Не следует удалять широколиственные растения. Известные места гнездования диких пчел могут быть отгорожены в качестве защиты от вспашки и затопления, но они не должны позволять подавиться растительностью. В этих областях следует отказаться от любого использования инсектицидов.
Щелочь Пчелиные гнездовья (Фото)
Исследования Др.Джордж Э. Бохарт из Министерство сельского хозяйства США обнаружило многие признаки дикой щелочи. пчелы для опыления, особенно в западной части Северной Америки. Рекомендации по их созданию указывает на климат с малым количеством осадков и бесструктурную почву минерального типа (песчаный или глиняный). Благоприятная местность это гамаки, невысокие гряды и пологие склоны, на которых никогда не останется воды поверхность земли. Подземный особенности — твердая сковорода или сковорода с естественным артезианским давлением.В некоторых случаях можно найти область, которая получает суб-орошение из поверхностных вод, подаваемых на орошаемые культуры на высший уровень. Но это может быть сложно поддерживать в стабильном состоянии. Участок должен быть рядом с полями, на которых выращиваются посевы, если только там нет никаких помех другим полям поблизости. На больших полях вокруг должно быть несколько гнездовий. Это. Щелочные пчелы кормят ближайших цветы сначала, а затем постепенно разлетаются.
Удаление растительности или как минимум прореживание до рассеянного рисунка должно начать подготовку к гнездованию места. Поверхностный дренаж может быть контролируется в низинных областях путем разбивки поверхности на серию низких гребни. Он должен быть защищен от паводковый полив и естественное половодье из ручьев. Почвенную влагу необходимо орошать в чтобы поддерживать почву во влажном состоянии до начала поверхности в начале лета и до осени.Влага должна присутствовать на глубине 5-10 дюймов. Если твердая сковорода уже присутствует, нужно проложить серию параллельных канав, чтобы уровень, расположенный так, чтобы области между ними были полностью пропитаны через несколько дни прохождения светового потока через канавы. Если твердой сковороды нет, ее можно создать, соскоблив 16-18 дюймов почвы и уплотнение грунта тяжелой техникой. Слой глины лучше подойдет результат. Растительность следует держать в минимум, потому что он расходует влагу.
Обслуживание сайтов очень важный. Густой рост сорняков должен быть предотвращенным. Дополнительное орошение часто бывает требуется для поддержания надлежащих условий влажности. Следует приложить усилия, чтобы получить достаточное количество влажность на поверхности во время сезона гнездования, чтобы предотвратить образование твердая щелочная корочка или сухой порошкообразный слой под тонкой корочкой черного цвета щелочь. Может понадобиться соскоблить от щелочи на поверхности с начала до конца лета.Сайт никогда не должен содержать стоячая вода. Скот может бродить место, но избегайте чрезмерного вытаптывания, особенно если поверхность имеет тенденцию высохнуть: оградить участок, если нужно. Скунсы и барсуки раскапывают личинки в любое время года и могут даже поймать взрослых особей. Их легко поймать или отравить. Пчелиные мухи, которые летают над норками с распростертыми крыльями бросают яйца во входы в гнезда и личинки уничтожают много личинок щелочных пчел.Усилия по контролировать их необходимо.
Больше гнездовых участков лучшее. Щелочные пчелы мигрируют и лучше всего стараться держать их рядом с желаемыми полями. Два или три незанятых участка поблизости помогут в их остающийся в районе.
Щелочные пчелы могут быть выведены естественная миграция, если поля находятся в пределах 1-2 миль от существующих участков.Пчелы также могут быть перемещены на участок. Личинки щелочных пчел можно использовать для ближние ходы. Блоки почвы с существующих участков могут быть пересажены. Стальные цилиндры можно вбивать в землю и содержащийся грунт. взолнованный. Взрослых можно поймать с помощью подметать сеть по мере их появления на существующем сайте. Из сети пчел можно пересыпать в бумажные мешки и поместите в прохладную влажную емкость. Их можно будет выпустить ближе к вечеру на предполагаемом гнездовье. места.
Экзамены & Упражнения
Ichneumonidae) в зависимости от количества яиц и опыта на участке — Университет Кюсю
TY — JOUR
T1 — Питание хозяином и принятие паразитической осой (Hymenoptera
T2 — Ichneumonidae) в зависимости от количества яиц и опыта патч
AU — Ueno, Takatoshi
N1 — Copyright: Авторское право 2004 г., Elsevier Science B.В., Амстердам. Все права защищены.
PY — 1999
Y1 — 1999
N2 — Оптимальные модели выбора хозяев, основанные на динамическом программировании, предсказывают, что физиологическое состояние насекомого-кормильца, то есть количество яиц, запасы энергии и т. Д., Влияет на поведенческие решения. Чтобы проверить это предсказание, было исследовано влияние физиологического состояния на принятие хозяином эктопаразитической осы Agrothereutes lanceolatus. Самкам ос в пластиковых стаканчиках (рассматриваемых как пятна) предъявляли хозяев, и их реакцию на хозяев постоянно наблюдали.После наблюдений осы препарировали и подсчитывали количество несенных ими зрелых и незрелых яиц. Результаты показали, что на поведенческие решения самок осы влияла нагрузка зрелых яиц, но не размер осы или количество незрелых яиц. Следовательно, осы с более высокой яйцекладкой с большей вероятностью откладывали яйца. Количество хозяев, ранее встречавшихся на участке (то есть опыт осы), также оказывало независимое влияние на принятие самками хозяев, указывая на то, что информационное состояние самок обновлялось во время поиска пищи на этом участке.Самки ос находились на кормлении только тогда, когда количество зрелых яиц приближалось к нулю. Одновременного кормления хозяев не наблюдалось. Выживаемость паразитоидов была почти нулевой, когда яйца паразитоидов переносились на хозяев, которых кормили, что указывает на то, что одновременное кормление хозяином могло вызвать высокую степень смертности потомства. Эти три результата подтвердили предположение и предсказание оптимальных моделей питания хозяев. Выбор паразитоидов-хозяев и кормление хозяев обсуждаются в контексте недавних моделей.
AB — Оптимальные модели выбора хозяев, основанные на динамическом программировании, предсказывают, что физиологическое состояние насекомого-кормильца, т.е.е. количество яиц, запасы энергии и т. д. влияют на поведенческие решения. Чтобы проверить это предсказание, было исследовано влияние физиологического состояния на принятие хозяином эктопаразитической осы Agrothereutes lanceolatus. Самкам ос в пластиковых стаканчиках (рассматриваемых как пятна) предъявляли хозяев, и их реакцию на хозяев постоянно наблюдали. После наблюдений осы препарировали и подсчитывали количество несенных ими зрелых и незрелых яиц. Результаты показали, что на поведенческие решения самок осы влияла нагрузка зрелых яиц, но не размер осы или количество незрелых яиц.Следовательно, осы с более высокой яйцекладкой с большей вероятностью откладывали яйца. Количество хозяев, ранее встречавшихся на участке (то есть опыт осы), также оказывало независимое влияние на принятие самками хозяев, указывая на то, что информационное состояние самок обновлялось во время поиска пищи на этом участке. Самки ос находились на кормлении только тогда, когда количество зрелых яиц приближалось к нулю. Одновременного кормления хозяев не наблюдалось. Выживаемость паразитоидов была почти нулевой, когда яйца паразитоидов переносились на хозяев, которых кормили, что указывает на то, что одновременное кормление хозяином могло вызвать высокую степень смертности потомства.Эти три результата подтвердили предположение и предсказание оптимальных моделей питания хозяев. Выбор паразитоидов-хозяев и кормление хозяев обсуждаются в контексте недавних моделей.
UR — http://www.scopus.com/inward/record.url?scp=0032766049&partnerID=8YFLogxK
UR — http://www.scopus.com/inward/citedby.url?scp=0032766049&partnerID=8YFLogx
U2 — 10.1023 / A: 1006582013488
DO — 10.1023 / A: 1006582013488
M3 — артикул
AN — SCOPUS: 0032766049
VL — 13
SP — 33
EP — 33 EP — эволюция ЭкологияJF — Эволюционная экология
SN — 0269-7653
IS — 1
ER —
Бактериальные ассоциаты семенных паразитических ос (Hymenoptera: Megastigmus) | BMC Microbiology
Schoonhoven LM, van Loon JJA, Dicke M: Insect-Plant Biology 2nd Edition. 2005, Oxford University Press, Нью-Йорк, США
Google Scholar
Янсон Е.М., Стироман Дж. О., Сингер М.С., Эббот П.: Мутуализмы насекомых-микробов-фитофагов и адаптивная эволюционная диверсификация. Эволюция. 2008, 62: 997-1012.
PubMed Google Scholar
Feldhaar H: Бактериальные симбионты как медиаторы экологически важных свойств насекомых-хозяев.Ecol Entomol. 2011, 36: 533-543.
Google Scholar
Диллон Р.Дж., Диллон В.М.: Кишечные бактерии насекомых: непатогенные взаимодействия. Анну Рев Энтомол. 2004, 49: 71-92.
CAS PubMed Google Scholar
Дуглас А.Э .: Микробное измерение в экологии питания насекомых. Funct Ecol. 2009, 23: 38-47.
Google Scholar
Энгель П., Моран Н.А.: Микробиота кишечника насекомых — разнообразие по структуре и функциям. FEMS Microbiol Rev.2013, 37: 699-735.
CAS PubMed Google Scholar
Накабачи А., Исикава Х: Обеспечение рибофлавином тли-хозяина, Acyrthosiphon pisum , эндосимбиотическими бактериями, Бухнера . J. Insect Physiol. 1999, 45: 1-6.
CAS PubMed Google Scholar
McCutcheon JP, Moran NA: Параллельная геномная эволюция и метаболическая взаимозависимость в древнем симбиозе. Proc Natl Acad Sci. 2007, 104: 19392-19397.
PubMed Central CAS PubMed Google Scholar
Warnecke F, Luginbhl P, Ivanova N, Ghassemian M, Richardson TH, Stege JT, Cayouette M, McHardy AC, Djordjevic G, Aboushadi N, Sorek R, Tringe SG, Podar M, Martin HG, Kunin V , Dalevi D, Madejska J, Kirton E, Platt D, Szeto E, Salamov A, Barry K, Михайлова N, Kyrpides NC, Matson EG, Ottesen EA, Zhang X, Hernndez M, Murillo C, Acosta LG: метагеномный и функциональный анализ микробиоты кишечника высших термитов, питающихся древесиной.Природа. 2007, 450: 560-565.
CAS PubMed Google Scholar
Уайтхед Л.Ф., Уилкинсон Т.Л., Дуглас А.Е.: Рециркуляция азота в симбиозе гороховой тли ( Acyrtosiphon pisum ). Proc R Soc London B Biol Sci. 1992, 250: 115-117.
Google Scholar
Adams AS, Aylward FO, Adams SM, Erbilgin N, Aukema BH, Currie CR, Suen G, Raffa KF: горные сосновые жуки, колонизирующие исторические и нефные деревья-хозяева, связаны с бактериальным сообществом, высоко обогащенным генами, способствующими к терпеновому метаболизму.Appl Environ Microbiol. 2013, 79: 3468-3475.
PubMed Central CAS PubMed Google Scholar
Гента Ф.А., Диллон Р.Дж., Терра В.Р., Феррейра С.Потенциальная роль кишечной микробиоты в переваривании клеточной стенки и детоксикации глюкозидов у личинок Tenebrio molitor . J. Insect Physiol. 2006, 52: 593-601.
CAS PubMed Google Scholar
Бауманн П. Биология эндосимбионтов, ассоциированных с бактериоцитами, насекомых, сосущих соки растений. Annu Rev Microbiol. 2005, 59: 155-189.
CAS PubMed Google Scholar
Моран Н.А., Маккатчеон Дж. П., Накабачи А: Геномика и эволюция наследственных бактериальных симбионтов. Анну Рев Жене. 2008, 42: 165-190.
CAS PubMed Google Scholar
Stouthamer R, Breeuwer JAJ, Hurst GDD: Wolbachia pipientis : микробный манипулятор размножения членистоногих.Annu Rev Microbiol. 1999, 53: 71-102.
CAS PubMed Google Scholar
Frago E, Dicke M, Godfray HCJ: Симбионты насекомых как скрытые игроки во взаимодействиях насекомых и растений. Trends Ecol Evol. 2012, 27: 705-711.
PubMed Google Scholar
Оливер К.М., Дегнан PH, Берк Г.Р., Моран Н.А.: Факультативные симбионты у тлей и горизонтальный перенос экологически важных признаков.Анну Рев Энтомол. 2010, 55: 247-266.
CAS PubMed Google Scholar
Цучида Т., Кога Р., Фукацу Т. Специализация растения-хозяина определяется факультативным симбионтом. Science 2004, 303: 1989.,
Генри Л.М., Пеккоуд Дж., Саймон Дж. К., Хэдфилд Дж. Д., Мэйден MJC, Феррари Дж., Годфрей HCJ: горизонтально передающиеся симбионты и колонизация экологических ниш хозяевами. Curr Biol. 2013, 23: 1-5.
Google Scholar
Брезнак Я.А.: Кишечная микробиота термитов и других насекомых-ксилофагов. Annu Rev Microbiol. 1982, 36: 323-343.
CAS PubMed Google Scholar
Bracke JW, Cruden DL, Markovetz AJ: Кишечная микробная флора американского таракана, Periplaneta americana L. Appl Environ Microbiol. 1979, 38: 945-955.
PubMed Central CAS PubMed Google Scholar
Глазго H: слепая кишка желудка и бактерии слепой кишки Heteroptera. Biol Bull. 1914, 26: 101-170.
Google Scholar
Джонс К.Г., Дауд П.Ф., Блэквелл М.: Полифилетическое происхождение дрожжевых эндоцитобионтов от анобиидных и церамбицидных жуков. Mycol Res. 1999, 103: 542-546.
Google Scholar
Налепа К.А., Бигнелл Д.Е., Банди С. Детритивори, копрофагия и эволюция пищеварительных мутуализмов у Dictyoptera.Insectes Soc. 2001, 48: 194-201.
Google Scholar
Хосокава Т., Кикучи Ю., Мэн XY, Фукацу Т.: Создание капсулы симбионта в жуке-вонючке платаспиде Megacopta punctatissima . FEMS Microbiol Ecol. 2005, 54: 471-477.
CAS PubMed Google Scholar
Shibata TF, Maeda T, Nikoh N, Yamaguchi K, Oshima K, Hattori M, Nishiyama T, Hasebe M, Fukatsu T, Kikuchi Y, Shigenobu S: Бактериальный симбионт бобового жука Riptortus pedestris .Объявление о геноме. 2013, 1: 1-2.
Google Scholar
Olivier-Espejel AS, Sabree ZL, Noge K, Becerra JX: В кишечной микробиоте нимфы и взрослых особей гигантского мескитового клопа ( Thasus neocalifornicus ) (Heteroptera: Coreidae) преобладает Burkholderia 4 de novo каждое поколение. Environ Entomol. 2011, 40: 1102-1110.
PubMed Google Scholar
Кикучи Ю., Хосокава Т., Фукацу Т.: Мутуализм насекомых-микробов без вертикальной передачи: каждое поколение насекомых-вонючих питает полезный симбионт из окружающей среды. Appl Environ Microbiol. 2007, 73: 4308-4316.
PubMed Central CAS PubMed Google Scholar
Хосокава Т., Кикучи Ю., Симада М., Фукацу Т.: Облигатный симбионт, участвующий в статусе вредителя насекомого-хозяина. Proc R Soc B Biol Sci. 2007, 274: 1979-1984.
CAS Google Scholar
Кукор Дж. Дж., Мартин М. М.: Получение пищеварительных ферментов сирицидными древесными отходами у их грибкового симбионта. Наука. 1983, 220: 1161-1163.
CAS PubMed Google Scholar
r? Tka P, Paoutov S, Kola? K M: Daldinia decipiens и Entonaema cinnabarina в качестве грибных симбионтов осы Xiphydria .Mycol Res. 2007, 111 (Pt 2): 224-231.
Google Scholar
Адамс А.С., Джордан М.С., Адамс С.М., Суен Дж., Гудвин Л.А., Дэвенпорт К.В., Карри С.Р., Раффа К.Ф.: бактерии, разрушающие целлюлозу, связанные с инвазивной мятой Sirex noctilio . ISME J. 2011, 5: 1323-1331.
PubMed Central CAS PubMed Google Scholar
Вебер Н.А.: Муравьи-грибники.Наука. 1966, 153: 587-604.
CAS PubMed Google Scholar
Мартинсон В.Г., Мой Дж., Моран Н.А.: Установление характерных кишечных бактерий во время развития пчеловодства. Appl Environ Microbiol. 2012, 78: 2830-2840.
PubMed Central CAS PubMed Google Scholar
Мартинсон В.Г., Данфорт Б.Н., Минкли Р.Л., Рюппелл О., Тингек С., Моран Н.А.: простая и характерная микробиота, связанная с медоносными пчелами и шмелями.Mol Ecol. 2011, 20: 619-628.
PubMed Google Scholar
Jeyaprakash A, Hoy MA, Allsopp MH: Бактериальное разнообразие взрослых особей рабочих особей Apis mellifera capensis и Apis mellifera scutellata (Insecta: Hymenoptera) оценено с использованием последовательностей 16S рРНК. J Invertebr Pathol. 2003, 84: 96-103.
CAS PubMed Google Scholar
Мор К.И., Теббе СС: Разнообразие и филотипическая консистенция бактерий в кишечнике трех видов пчел (Apoidea) на поле масличного рапса. Environ Microbiol. 2006, 8: 258-272.
CAS PubMed Google Scholar
Olofsson TC, Vsquez A: Обнаружение и идентификация новой молочнокислой бактериальной флоры в медовом желудке медоносной пчелы Apis mellifera . Curr Microbiol. 2008, 57: 356-363.
CAS PubMed Google Scholar
Моран Н.А., Хансен А.К., Пауэлл Дж. Э., Сабри З. Л.: Отличительная кишечная микробиота медоносных пчел, оцененная с помощью глубокого отбора проб у отдельных рабочих пчел. PLoS One 2012, 7: e36393.,
Van Borm S, Buschinger A, Boomsma JJ, Billen J: Tetraponera муравьи имеют кишечные симбионты, связанные с азотфиксирующими корневыми клубеньковыми бактериями. Proc R Soc London B Biol Sci. 2002, 269: 2023-2027.
CAS Google Scholar
Андерсон К.Э., Рассел Д.А., Моро С.С., Каутц С., Саллам К.Э., Ху Й., Бейсингер Ю., Мотт Б.М., Бак Н., Уиллер Д.Э .: Очень похожие микробные сообщества характерны для родственных и трофически схожих видов муравьев. Mol Ecol. 2012, 21: 2282-2296.
PubMed Google Scholar
Рассел Дж. А., Моро К. С., Гольдман-Уэртас Б., Фудзивара М., Ломан Д. Д., Пирс Н. Э.: Бактериальные симбионты кишечника тесно связаны с эволюцией травоядных животных у муравьев.Proc Natl Acad Sci. 2009, 106: 21236-21241.
PubMed Central CAS PubMed Google Scholar
Дегнан П., Лазарус А., Брок С., Вернегрин Дж. Стабильность симбионта-хозяина и быстрые темпы эволюции в ассоциации муравьев и бактерий: совместное видообразование видов Camponotus и их эндосимбионтов, Candidatus Blochmannia . Syst Biol. 2004, 53: 95-110.
PubMed Google Scholar
Gil R, Silva FJ, Zientz E, Delmotte F, Gonzlez-Candelas F, Latorre A, Rausell C, Kamerbeek J, Gadau J, Hlldobler B, van Ham RCHJ, Gross R, Moya A: последовательность генома blochmannia floridanus : сравнительный анализ редуцированных геномов. Proc Natl Acad Sci. 2003, 100: 9388-9393.
PubMed Central CAS PubMed Google Scholar
Degnan PH, Lazarus AB, Wernegreen JJ: Последовательность генома Blochmannia pennsylvanicus указывает на параллельные эволюционные тенденции среди бактериальных мутуалистов насекомых.Genome Res. 2005, 15: 1023-1033.
PubMed Central CAS PubMed Google Scholar
Feldhaar H, Straka J, Krischke M, Berthold K, Stoll S, Mueller MJ, Gross R: Улучшение питания всеядных муравьев-плотников эндосимбионтом Blochmannia . BMC Biol 2007, 5:48.,
Giron D, Kaiser W., Imbault N, Casas J: Цитокинин-опосредованное манипулирование листом гусеницей минера.Biol Lett. 2007, 3: 340-343.
PubMed Central CAS PubMed Google Scholar
Giron D, Frago E, Glevarec G, Pieterse CMJ, Dicke M: Цитокинины как ключевые регуляторы во взаимодействиях растений, микробов и насекомых: соединение роста растений и защиты. Funct Ecol. 2013, 27: 599-609.
Google Scholar
Стоун Г. Н., Шнрогге К. Адаптивное значение морфологии галлов насекомых.Trends Ecol Evol. 2003, 18: 512-522.
Google Scholar
Schwachtje J, Baldwin IT: Почему атака травоядных изменяет первичный метаболизм ?. Plant Physiol. 2008, 146: 845-851.
PubMed Central CAS PubMed Google Scholar
Kaiser W, Huguet E, Casas J, Commin C, Giron D: Фенотип зеленого острова растений, индуцированный листовыми минерами, опосредуется бактериальными симбионтами.Proc R Soc B Biol Sci. 2010, 277: 2311-2319.
CAS Google Scholar
Джеймсон П: Цитокинины и ауксины во взаимодействиях между растениями и патогенами Обзор. Регул роста растений. 2000, 32: 369-380.
CAS Google Scholar
Сакакибара H: Цитокинины: активность, биосинтез и транслокация. Annu Rev Plant Biol. 2006, 57: 431-449.
CAS PubMed Google Scholar
Frugier F, Kosuta S, Murray JD, Crespi M, Szczyglowski K: Цитокинин: секретный агент симбиоза. Trends Plant Sci. 2008, 13: 115-120.
CAS PubMed Google Scholar
Body M, Kaiser W, Dubreuil G, Casas J, Giron D: Минеры кооптируют микроорганизмы для улучшения своей питательной среды. J Chem Ecol. 2013, 39: 969-977.
CAS PubMed Google Scholar
Auger-Rozenberg M.-A, Roques A: Инвазии семенных ос, вызванные нерегулируемой торговлей семенами, влияют на биоразнообразие растений и животных. Интегр Зоол. 2012, 7: 228-246.
PubMed Google Scholar
Grissell EE: Аннотированный каталог мировых мегастигминов (Hymenoptera: Chalcidoidea: Torymidae). Contrib Am Entomol Inst. 1999, 31: 1-92.
Google Scholar
Roques A, Skrzypczyμska M: Заражающие семена хальциды из рода Megastigmus Dalman, 1820 (Hymenoptera: Torymidae), родом из Западного Палеарктического региона: таксономия, специфичность хозяев и распространение. J Nat Hist. 2003, 37: 127-238.
Google Scholar
фон Адеркас П., Руо Г., Вагнер Р., Рор Р., Рокес А: Паразитизм семян перенаправляет развитие семяпочек у пихты Дугласовой. Proc R Soc B Biol Sci. 2005, 272: 1491-1496.
Google Scholar
фон Aderkas P, Rouault G, Wagner R, Chiwocha S, Roques A: Многоядерные накопительные клетки в пихте Дугласа ( Pseudotsuga menziesii (Mirbel) Franco) и эффект паразитизма семян, вызванный хальцидом spermotrophus Wachtl. Наследственность. 2005, 94: 616-622.
CAS PubMed Google Scholar
Chiwocha S, Rouault G, Abrams S, von Aderkas P: Паразитизм семян пихты Дугласа ( Pseudotsuga menziesii ) семенами хальцида, Megastigmus spermotrophus и его влияние на физиологию семенного гормона.Половое растение Reprod. 2007, 20: 19-25.
CAS Google Scholar
Rouault G, Turgeon J, Candau J-N, Roques A, von Aderkas P: Стратегии откладки яиц хальцидов из семян хвойных пород в зависимости от фенологии хозяина. Naturwissenschaften. 2004, 91: 472-480.
CAS PubMed Google Scholar
Дурон О., Бушон Д., Бутин С., Беллами Л., Чжоу Л., Энгельстдтер Дж., Херст Г. Д.: Разнообразие репродуктивных паразитов среди членистоногих: Wolbachia не ходят одни. BMC Biol 2008, 6:27.,
Boivin T, Henri H, Vaver F, Gidoin C, Candau JN, Magnoux E, Roques A, Auger-Rozenbert MA: Эпидемиология телитоки, вызванная эндосимбиотиком. Wolbachia среди видов ос, специализирующихся на семенах: специализация растений-хозяев имеет значение. Mol Ecol. 2014, 23: 2362-2375.
CAS PubMed Google Scholar
Вайнерт Л.А., Веррен Дж. Х., Эби А., Стоун Г. Н., Джиггинс Ф. М.: Эволюция и разнообразие бактерий Rickettsia . BMC Biol 2009, 7: 6.,
Ludwig W, Schleifer K-H, Whitman WB: Пересмотренная дорожная карта к типу Firmicutes. Bergeys Manual of Systematic Bacteriology Vol 3. 2008, Springer, New York
Google Scholar
Цуг Р., Хаммерштейн П.: Все еще множество хозяев для Wolbachia : анализ последних данных показывает, что 40% наземных видов членистоногих инфицированы. PLoS One 2012, 7: e38544.,
Werren JH, Baldo L, Clark ME: Wolbachia : мастера-манипуляторы биологии беспозвоночных. Nat Rev Microbiol. 2008, 6: 741-751.
CAS PubMed Google Scholar
Stouthamer R: Wolbachia -индуцированный партеногенез. Влиятельные пассажиры. Под редакцией: ONeill SL, Werren JH. 1997, Oxford University Press, Нью-Йорк, США, 102–124.
Google Scholar
Rokas A, Atkinson RJ, Nieves-Aldrey J-L, West SA, Stone GN: Распространенность и разнообразие Wolbachia в желчнокаховых (Hymenoptera; Cynipidae) на дубе. Mol Ecol. 2002, 11: 1815-1829.
CAS PubMed Google Scholar
Plantard O, Rasplus J, Clainche Le I, Solignac M: Wolbachia -индуцированная телитокия в желчнокаменной желчнице Diplolepis spinosissimae (Giraud) (Hymenoptera: Cynipidae) и его генетические последствия. его хозяин.Proc R Soc London B Biol Sci. 1998, 265: 1075-1080.
Google Scholar
Перлман С.Дж., Хантер М.С., Жори-Фейн Э.: Возникающее разнообразие Риккетсий . Proc R Soc B Biol Sci. 2006, 273: 2097-2106.
Google Scholar
Werren JH, Hurst GD, Zhang W, Breeuwer JA, Stouthamer R, Majerus ME: риккетсиозный родственник, связанный с убийством самцов жука божьей коровки ( Adalia bipunctata ).J Bacteriol. 1994, 176: 388-394.
PubMed Central CAS PubMed Google Scholar
Хагимори Т., Абэ Й., Дате С., Миура К.: первая находка бактерии Rickettsia , связанной с индукцией партеногенеза среди насекомых. Curr Microbiol. 2006, 52: 97-101.
CAS PubMed Google Scholar
Тейшейра Л., Феррейра А., Эшбёрнер М.: Бактериальный симбионт Wolbachia индуцирует устойчивость к вирусным инфекциям РНК у Drosophila melanogaster . PLoS Biol 2008, 6: e2.,
? Ukasik P, van Asch M, Guo H, Ferrari J, Godfray HCJ: Неродственные факультативные эндосимбионты защищают тлю от грибкового патогена. Ecol Lett. 2013, 16: 214-218.
PubMed Google Scholar
Герер Л., Ворбургер С. Паразитоиды как переносчики факультативных бактериальных эндосимбионтов у тлей. Biol Lett. 2012, 8: 613-615.
PubMed Central PubMed Google Scholar
Duron O, Wilkes TE, Hurst GDD: Межвидовая передача бактерии, убивающей самцов, в экологической временной шкале. Ecol Lett. 2010, 13: 1139-1148.
PubMed Google Scholar
Брессан А., Смти О., Арнеодо Дж., Лерминье Дж., Будон-Падье Э: Вектор передачи патогенной бактерии в кладе Arsenophonus , разделяющей экологические черты с факультативными эндосимбионтами насекомых. Фитопатология. 2009, 99: 1289-1296.
CAS PubMed Google Scholar
Яенике Дж., Полак М., Фискин А., Хелу М., Минхас М.: Межвидовая передача эндосимбиотической Spiroplasma клещами. Biol Lett. 2007, 3: 23-25.
PubMed Central CAS PubMed Google Scholar
Моран Н.А., Данбар Х.Э .: Получение полезных симбионтов половым путем у тлей. Proc Natl Acad Sci U S A.2006, 103: 12803-12806.
PubMed Central CAS PubMed Google Scholar
Gatineau F, Jacob N, Vautrin S, Larrue J, Lherminier J, Richard-Molard M, Boudon-Padieu E: Связь с синдромом Басса. Богатство сахарной свеклы передается фитоплазмой и бактериоподобным организмом с помощью Pentastiridius sp. Фитопатология. 2002, 92: 384-392.
CAS PubMed Google Scholar
Danet J, Foissac X, Zreik L, Salar P, Verdin E, Nourrisseau J, Garnier M: Candidatus Phlomobacter fragariae — распространенный возбудитель маргинального хлороза клубники на производственных полях во Франции и передается через цикадку Cixius wagneri (Китай). Фитопатология. 2002, 93: 644-649.
Google Scholar
Colman DR, Toolson EC, Takacs-Vesbach CD: Влияют ли диета и таксономия на бактериальные сообщества кишечника насекомых ?.Mol Ecol. 2012, 20: 5124-5137.
Google Scholar
Brucker RM, Bordenstein SR: Роли эволюционных отношений хозяина (род: Nasonia ) и развитие в структурировании микробных сообществ. Эволюция. 2012, 66: 349-362.
PubMed Google Scholar
Каутц С., Рубин Б.Э., Рассел Дж. А., Моро К.С.: Изучение микробиома муравьев: сравнение пиросеквенирования 454 с традиционными методами для выявления бактериального разнообразия.Appl Environ Microbiol. 2013, 79: 525-534.
PubMed Central CAS PubMed Google Scholar
Ishak HD, Ploughes R, Sen R, Kellner K, Meyer E, Estrada DA, Dowd SE, Mueller UG: Бактериальное разнообразие в Solenopsis invicta и Solenopsis geminata колоний муравьев, характеризующихся ампликоном 16S пипрозы 454 . Microb Ecol. 2011, 61: 821-831.
PubMed Google Scholar
Janssen PH: Определение доминирующих таксонов почвенных бактерий в библиотеках генов 16S рРНК и 16S рРНК. Appl Environ Microbiol. 2006, 72: 1719-1728.
PubMed Central CAS PubMed Google Scholar
Да Роча ООН, ван Овербек Л., ван Эльзас Дж. Д.: Изучение ранее не культивируемых бактерий из ризосферы. FEMS Microbiol Ecol. 2009, 69: 313-328.
PubMed Google Scholar
Mundt JO, Hinkle NF: Бактерии в семязачатках и семенах. Appl Environ Microbiol. 1976, 32: 694-698.
PubMed Central CAS PubMed Google Scholar
Mukhopadhyay K, Garrison NK, Hinton DM, Bacon CW, Khush GS, Peck HD, Datta N: Идентификация и характеристика бактериальных эндофитов риса. Микопатология. 1996, 134: 151-159.
CAS PubMed Google Scholar
Hallmann J, Quadt-Hallmann A, Mahaffee WF, Kloepper JW: Бактериальные эндофиты сельскохозяйственных культур. Может J Microbiol. 1997, 43: 895-914.
CAS Google Scholar
Шарки MJ: Филогения и классификация перепончатокрылых. Zootaxa. 2007, 1668: 521-548.
Google Scholar
Хаким Р.С., Болдуин К., Смагге Г.: Регулирование роста, развития и метаморфоза средней кишки.Анну Рев Энтомол. 2010, 55: 593-608.
CAS PubMed Google Scholar
Bution ML, Caetano FH: Симбиотические бактерии и структурные особенности подвздошной кишки Cephalotes муравьев. Микрон. 2010, 41: 373-381.
PubMed Google Scholar
Бреннер Д. Д., Криг Н. Р., Стейли Дж. Т.: Том 2, часть C, Протеобактерии; альфа-, бета-, дельта- и эпсилонпротеобактерии.Руководство Бергейса по систематической бактериологии. 2005, Спрингер, Нью-Йорк, 2
Google Scholar
Джонс Р.Т., Маккормик К.Ф., Мартин А.П.: Бактериальные сообщества блох с положительной реакцией на Bartonella : разнообразие и схемы сборки сообществ. Appl Environ Microbiol. 2008, 74: 1667-1670.
PubMed Central CAS PubMed Google Scholar
Lundgren JG, Lehman RM: Бактериальные симбионты кишечника способствуют перевариванию семян всеядного жука. PLoS One 2010, 5: e10831.,
Hail D, Dowd SE, Bextine B: Идентификация и расположение симбионтов, связанных с картофельным листоносом ( Bactericera cockerelli ) Жизненные этапы. Environ Entomol. 2012, 41: 98-107.
PubMed Google Scholar
Husnik F, Nikoh N, Koga R, Ross L, Duncan RP, Fujie M, Tanaka M, Satoh N, Bachtrog D, Wilson ACC, von Dohlen CD, Fukatsu T, McCutcheon JP: горизонтальный перенос генов из Разнообразие бактерий в геноме насекомого обеспечивает трехсторонний симбиоз гнездового мучнистого червеца.Клетка. 2013, 153: 1567-1578.
CAS PubMed Google Scholar
Vaneechoutte M, Kmpfer P, Thierry DB, Falsen E, Verschraegen G: Wautersia gen. nov., новый род, включающий филогенетическую линию, включающую Ralstonia eutropha и родственные виды, и предложение Ralstonia [ Pseudomonas ] syzygii (Roberts et al. 1990) comb. ноя Int J Syst Evol Microbiol.2004, 54: 317-327.
PubMed Google Scholar
Кикучи Ю., Хосокава Т., Фукацу Т.: Древняя, но беспорядочная ассоциация хозяин-симбионт между кишечными симбионтами Burkholderia и их гетероптеранными хозяевами. ISME J. 2011, 5: 446-460.
PubMed Central PubMed Google Scholar
Кикучи Ю., Мэн Х, Фукацу Т: Кишечные симбиотические бактерии рода Burkholderia у широкоголовых клопов Riptortus clavatus и Leptocorisa chinensis (Heteroptera): Alydidaera.Appl Environ Microbiol. 2005, 71: 4035-4043.
PubMed Central CAS PubMed Google Scholar
Stone SL, Gifford DJ: Структурные и биохимические изменения в семенах сосны лоблолли (Pinus taeda L.) во время прорастания и раннего роста проростков. II. Хранение триацилглицеринов и углеводов. Int J Plant Sci. 1999, 160: 663-671.
CAS Google Scholar
King JE, Gifford DJ: Использование аминокислот в семенах сосны лоблолли во время прорастания и раннего роста проростков. 1. Аргинин и активность аргиназы. Plant Physiol. 1997, 113: 1125-1135.
PubMed Central CAS PubMed Google Scholar
Тодд С.Д., Гиффорд Д.Д.: Роль мегагаметофита в поддержании экспрессии гена аргиназы в проростках сосны лоблолли ( Pinus taeda L.) in vitro. Planta. 2002, 215: 110-118.
CAS PubMed Google Scholar
Pant R: Выведение азота насекомыми. Proc Anim Sci. 1988, 97: 379-415.
CAS Google Scholar
Rosenthal GA, Janzen DH, Dahlman DL, Url S, Carolina N, Hill C: деградация и детоксикация канаванина специализированным семенным хищником. Наука. 1977, 196: 658-660.
CAS PubMed Google Scholar
Сабри З.Л., Камбхампати С., Моран Н.А.: Рециркуляция азота и обеспечение питательными веществами Blattabacterium , эндосимбионта тараканов. Proc Natl Acad Sci. 2009, 106: 19521-19526.
PubMed Central CAS PubMed Google Scholar
Zientz E, Dandekar T, Gross R: Метаболическая взаимозависимость облигатных внутриклеточных бактерий и их насекомых-хозяев. Microbiol Mol Biol Rev.2004, 68: 745-770.
PubMed Central CAS PubMed Google Scholar
Кашима Т., Накамура Т., Тодзё С: рециркуляция мочевой кислоты в щитовидном клопе, Parastrachia japonensis (Hemiptera: Parastrachiidae), во время диапаузы. J. Insect Physiol. 2006, 52: 816-825.
CAS PubMed Google Scholar
Чен В.М., Джеймс Э.К., Прескотт А.Р., Киранс М., Спрент Д.И.: Нодуляция Mimosa spp. бета-протеобактериями Ralstonia taiwanensis . Мол Растительный Микроб Взаимодействовать.2003, 16: 1051-1061.
CAS PubMed Google Scholar
Тамура К., Петерсон Д., Петерсон Н., Стечер Г., Ней М., Кумар С.: MEGA5: Анализ молекулярной эволюционной генетики с использованием методов максимального правдоподобия, эволюционного расстояния и максимальной экономии. Mol Biol Evol. 2011, 28: 2731-2739.
PubMed Central CAS PubMed Google Scholar
Wang H-X, Geng Z-L, Zeng Y, Shen Y-M: обогащение микробиоты растений для создания метагеномной библиотеки.Environ Microbiol. 2008, 10: 2684-2691.
CAS PubMed Google Scholar
Айрес Т., Марб Н., Серрао Э.А., Дуарте С.М., Арно-Хаонд С. Селективное удаление хлоропластидной ДНК для метагеномики бактерий, связанных с зелеными водорослями Caulerpa taxifolia (Bryopsidophyceae). J Phycol. 2012, 48: 483-490.
Google Scholar
Palavesam A, Guerrero FD, Heekin AM, Wang J, Dowd SE, Sun Y, Foil LD, de Prez Len AA: анализ микробиома, ассоциированного с Horn Fly, на основе пиросеквенирования, Haematobia раздражает . PLoS One 2012, 7: e44390.,
Shange RS, Ankumah RO, Ibekwe AM, Zabawa R, Dowd SE: отчетливые сообщества почвенных бактерий, выявленные в агроэкосистеме с различным управлением. PLoS One 2012, 7: e40338.,
Dowd SE, Callaway TR, Wolcott RD, Sun Y, McKeehan T, Hagevoort RG, Edrington TS: Оценка бактериального разнообразия в фекалиях крупного рогатого скота с использованием 16S Пиросеквенирование ампликона FLX, кодируемого рДНК, бактериальной меткой (bTEFAP). BMC Microbiol 2008, 8: 125.,
Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, Lesniewski RA, Oakley BB, Parks DH, Robinson CJ, Sahl JW, Stres B, Thallinger GG, Van Horn DJ, Вебер CF: Представляем mothur: программное обеспечение с открытым исходным кодом, независимое от платформы, поддерживаемое сообществом для описания и сравнения сообществ микробов. Appl Environ Microbiol. 2009, 75: 7537-7541.
PubMed Central CAS PubMed Google Scholar
Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, Fierer N, Pea AG, Goodrich JK, Gordon JI, Huttley GA, Kelley ST, Knights D, Koenig JE, Ley RE, Lozupone CA, Mcdonald D, Muegge BD, Pirrung M, Reeder J, Sevinsky JR, Turnbaugh PJ, Walters WA, Widmann J, Yatsunenko T., Zaneveld J, Knight R: QIIME позволяет анализировать высокопроизводительные данные секвенирования сообщества. Нат методы. 2010, 7: 335-336.
PubMed Central CAS PubMed Google Scholar
Ридер Дж., Найт Р.: Быстрое устранение шумов в считывании ампликонов пиросеквенирования путем использования распределений ранг-распространенность. Нат методы. 2010, 7: 668-669.
PubMed Central CAS PubMed Google Scholar
Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R: UCHIME улучшает чувствительность и скорость обнаружения химер. Биоинформатика. 2011, 27: 2194-2200.
PubMed Central CAS PubMed Google Scholar
Edgar RC: поиск и кластеризация на порядки быстрее, чем BLAST. Биоинформатика. 2010, 26: 2460-2461.
CAS PubMed Google Scholar
Ван К., Гаррити Г.М., Тидье Дж. М., Коул Дж. Р.: Байесовский классификатор Нава для быстрого отнесения последовательностей рРНК к новой бактериальной таксономии. Appl Environ Microbiol. 2007, 73: 5261-5267.
PubMed Central CAS PubMed Google Scholar
Макдональд Д., Прайс М.Н., Гудрич Дж., Навроцки Е.П., ДеСантис Т.З., Пробст А., Андерсен Г.Л., Найт Р., Гугенгольц П.: улучшенная таксономия Грингена с явными рангами для экологического и эволюционного анализа бактерий и архей. ISME J. 2012, 6: 610-618.
PubMed Central CAS PubMed Google Scholar
Вернер Дж., Корен О., Хугенгольц П., ДеСантис Т.З., Уолтерс В.А., Капорасо Дж. Г., Ангенент Л.Т., Найт Р., Лей Р.Э .: Влияние обучающих наборов на классификацию высокопроизводительных исследований генов 16s рРНК бактерий.ISME J. 2012, 6: 94-103.
PubMed Central CAS PubMed Google Scholar
Nawrocki EP, Kolbe DL, Eddy SR: Infernal 1.0: вывод выравнивания РНК. Биоинформатика. 2009, 25: 1335-1337.
PubMed Central CAS PubMed Google Scholar
Прайс М.Н., Дехал П.С., Аркин А.П.: FastTree 2 — деревья приблизительно максимального правдоподобия для больших трасс. PLoS One 2010, 5: e9490.,
Sourial N, Wolfson C, Zhu B, Quail J, Fletcher J, Karunananthan S, Bandeen-Roche K, Bland F, Bergman H: Анализ соответствия представляет собой полезный инструмент для выявления взаимосвязей между категориальными переменными. J Clin Epidemiol. 2010, 63: 638-646.
PubMed Central PubMed Google Scholar
Язык и среда для статистических вычислений. 2013, Фонд R для статистических вычислений, Вена, Австрия
Макмурди П.Дж., Холмс С.Филосек: пакет биопроводников для обработки и анализа данных филогенетической последовательности с высокой пропускной способностью. Pacific Symp Biocomput. 2012, 17: 235-246.
Google Scholar
Dray S, Dufour A-B: Пакет ade4: Реализация диаграммы двойственности для экологов. J Stat Softw. 2007, 22: 1-20.
Google Scholar
Lozupone C, Knight R: UniFrac: новый филогенетический метод сравнения микробных сообществ.Appl Environ Microbiol. 2005, 71: 8228-8235.
PubMed Central CAS PubMed Google Scholar
Hamady M, Lozupone C, Knight R: Fast UniFrac: обеспечение высокопроизводительного филогенетического анализа микробных сообществ, включая анализ данных пиросеквенирования и PhyloChip. ISME J. 2010, 4: 17-27.
PubMed Central CAS PubMed Google Scholar
Marchesi JR, Sato T, Weightman AJ, Martin TA, Fry JC, Hiom SJ, Dymock D, Wade WG: Дизайн и оценка полезных бактериально-специфичных праймеров для ПЦР, которые амплифицируют гены, кодирующие бактериальную 16S рРНК.
